
1. Docente, Universidad del Tolima, Facultad de Ciencias, Ibagué. Investigador Asociado, Laboratorio de Biología Molecular y Patogénesis, Universidad del Valle, Cali.
2. Profesor Titular, Departamento de Ciencias Fisiológicas, Facultad de Salud. Director, Laboratorio de Biología Molecular y Patogénesis, Universidad del Valle, Cali.
RESUMENSe examinó la integración del virus HTLV-I en 16 personas seropositivas; 11 pacientes con PET/HAM, 2 que presentaban otras enfermedades y 3 asintomáticos. El ADN de los linfocitos de la sangre periférica se sometió a una prueba de IPCR y los resultados obtenidos mostraron evidencias de una integración y patrones de expansión oligoclonal para el HTLV-I. Además se observó una tendencia hacia un número mayor de integraciones entre las personas con PET/HAM (7.2 ± 2) en contraste con los que padecen enfermedades diferentes (4 ± 1) o con los individuos asintomáticos incluidos en el estudio (4 ± 2.2). Se determinó que los pacientes con PET/HAM tienen subpoblaciones de células infectadas con HTLV-I que han proliferado de manera oligoclonal. Los resultados obtenidos mediante amplificación de las secuencias flanqueantes al provirus sugieren la existencia de preferencialidad en los sitios de integración del provirus HTLV-I en células linfocitarias de los pacientes con PET/HAM.
El virus linfotrópico humano tipo I (HTLV-I) es un retrovirus incluido dentro del grupo HTLV, STLV, BLV de la subfamilia Oncovirinae. Se asocia, sobre todo, con la leucemia de células T del adulto (ATL, por las siglas en inglés)1-5 y la paraparesia espástica tropical/mielopatía asociada con la infección por HTLV-I (PET/HAM)6-21, aunque cada día son más fuertes las asociaciones con otras entidades clínicas como uveítis, artropatías, dermatitis infecciosa y la polimiositis16. El HTLV-I es endémico en diversas regiones geográficas que incluyen Sur y Norteamérica, el sudeste de Japón, la región del Caribe y áfrica. Como en muchas otras regiones, el virus se restringe geográficamente. En Colombia, el municipio suroccidental de Tumaco, sobre el Océano Pacífico, es una zona endémica para el HTLV-I, pues se ha caracterizado por una elevada relación en el aumento anual de la PET/HAM17.
Se han visto secuencias no específicas con determinados resultados clínicos de la enfermedad; mientras tanto, en estudios moleculares, se demostró que en algunas partes del genoma del HTLV-I ciertos cambios en la secuencia nucleotídica se correlacionan con el origen geográfico de los pacientes22. Los factores responsables del progreso de infección a enfermedad todavía no se entienden por completo, aunque la activación de las células linfoides en el huésped puede aumentar la expresión de genes virales y su replicación. La producción de bandas monorreactivas IgG e IgA por parte de los linfocitos B contra una proteína híbrida recombinante gag-env, aumenta de modo significativo en pacientes con PET/HAM en contraste con los portadores asintomáticos y los individuos seronegativos para el virus23.
El HTLV-I se integra en el genoma de los pacientes con ATL y de los afectados por PET/HAM y según la línea de evidencias que se acepta tradicionalmente, se cree que el proceso de integración en el genoma del huésped humano ocurre al azar. Más recientemente se ha consolidado una segunda línea de evidencias menos conocida, fundamentada en las observaciones hechas con base en el fraccionamiento en la composición del genoma humano; según las mismas, existe una preferencialidad de sitios de integración en los isocoros ricos en GC24,25; asimismo, mediante clonación y secuenciación de la región genómica flanqueante al provirus HTLV-I se observa que ésta es rica en AT, lo que sugiere sitios de integración preferencial dentro del genoma de individuos infectados5,33.
En el desarrollo de la ATL se requiere la integración previa del virus26, que se efectúa de un modo monoclonal27 y se ha sugerido que este patrón clonal de integración es un indicador biológico particular del estado leucémico. Estos hechos proporcionan evidencias directas de la participación del virus en el desarrollo de la ATL. Por otra parte, algunas evidencias polémicas informan la existencia de una correlación entre los patrones de integración y el curso clínico de la ATL28,29.
En pacientes con PET/HAM los sitios de integración del HTLV-I varían30, 31, se localizan en distintas regiones cromosómicas y se ha informado que su modo de integración es policlonal aunque también de forma aleatoria26,29,33,43,45, tanto en portadores asintomáticos como en individuos con PET/HAM, en contraste con lo que sucede en la ATL.
Mediante técnicas basadas en la reacción en cadena de la polimerasa (PCR, siglas en inglés) se ha demostrado la expansión clonal de las células infectadas por HTLV-I de pacientes con PET/HAM en diversos estadíos de la enfermedad34,35. En este trabajo se analizó mediante la técnica de PCR inversa la modalidad de integración del HTLV-I en células infectadas in vivo, lo mismo que la variación de los sitios de integración del HTLV-I en pacientes con PET/HAM.
MATERIALES Y MéTODOS
La muestra. Se incluyeron 16 individuos seropositivos por ELISA, Western Blot y PCR para HTLV-I. De ellos, 11 tenían paraparesia espástica tropical, 3 eran portadores asintomáticos y 2 estaban afectados con enfermedades distintas. Además, los pacientes PET/HAM seropositivos para HTLV-I provenían de diversos sitios de la zona pacífica en Colombia.
Extracción del ADN de linfocitos de pacientes seropositivos para HTLV-I. Los mononucleares periféricos se aislaron de las muestras sanguíneas mediante separación en gradientes de Ficoll-Hipaque (Histopaque®), se lavaron dos veces en amortiguador fosfato salino (PBS)36 y el precipitado se lisó inmediatamente por tratamiento con SDS/proteinasa K a 56º C. El ADN se extrajo por precipitación en etanol y fenol, y el precipitado se resuspendió en 10 mmol/l de Tris-HCL pH 8.0, 1 mmol/l de EDTA36. La concentración y el grado de pureza se midieron por fluorimetría. Se tomaron las precauciones necesarias para evitar la contaminación en el área donde se hizo la extracción del ADN.
Amplificación del ADN genómico flanqueante al provirus HTLV-I por PCR inversa. Para llevar a cabo la amplificación enzimática mediante la técnica de PCR inversa (IPCR) se utilizó la modificación del protocolo desarrollado por Take-moto et al.37, y puesto en marcha en 1998 en el Laboratorio de Biología Molecular y Patogénesis de la Universidad del Valle 38. El proceso comprende una etapa de digestión con la enzima de restricción Alu I, una autoligación y posterior digestión de los anillos formados por Sst II36. Finalmente, los autoligados no digeridos por Sst II se amplificaron por PCR (Figura 1).
Como cebadores de la PCR se emplearon el HTVL-009, 5´-AAGCCGGCAGTCAGTCGTGA-3´ (8946-8927 nt) y el HTLV-010, 5´-AAGTACCGGCAACTCTGCTG-3´ (8958-8977). Previamente a la realización de los ciclos de reacciones de PCR se sometieron las mezclas a una temperatura de 93º C durante 30 minutos para relajar el ADN circular y permitir su amplificación. La reacción de alineación se hizo a 42º C durante 30 segundos y la de extensión a 72º C por 30 segundos. Se efectuaron 40 ciclos de amplificación en un termociclador Cetus Perkin Elmer. Los productos de la PCR se visualizaron mediante electroforesis en geles de agarosa al 2% con bromuro de etidio y para aumentar la resolución en geles de poliacrilamida por tinción con plata36. Se hizo un análisis para determinar el número de amplicones por recuento según el número de bandas por muestra (un amplicón significó expansión monoclonal; más de un amplicón, oligoclonal).
RESULTADOS
De las 16 muestras de PBMC de pacientes seropositivos infectados por el HTLV-I todos mostraron amplificación de segmentos de ADN por IPCR. Sin embargo, el número de amplicones registrado fue variable tanto en tamaño como en la distribución de ellos (Figura 2).
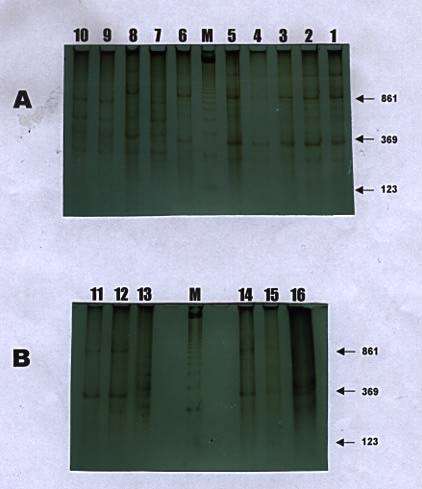
Un análisis detallado del tamaño y la frecuencia de cada amplicón en los pacientes con PET/HAM mostró que existía una distribución diferencial en la que algunos amplicones estaban más representados que otros (Cuadro 1).


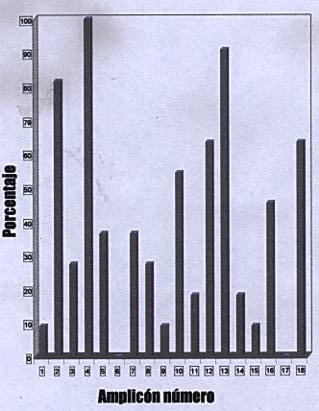
El amplicón de 230 pb mostró una frecuencia de 81.3% (13/16), el de 300 pb está representado en 100% (16/16) y el de 860 pb se determinó en su frecuencia de 81.3% (13/16). Para los amplicones de 300 y 860 pb que presentan 81.3% no todos los individuos tenían simultáneamente el mismo patrón de bandas. El resto de amplicones mostró en general una baja frecuencia y coamplificación variable.
Uno de los aspectos más importantes del estudio era analizar la distribución de amplicones IPCR de manera exclusiva en enfermos con PET/HAM. Para tal efecto, se incluyeron 11 pacientes PET/HAM, en quienes se obtuvo un número promedio de amplicones de 7.2 ± 2, en contraste con el de los asintomáticos que fue 4 ± 2.2 y el promedio de 4 ± 1 en los de otras enfermedades.
Un análisis detallado de la distribución de amplicones en el total de los 11 individuos PET/HAM reveló la presencia de un patrón preferencial de distribución de amplicones. La coamplificación simultánea de los fragmentos de IPCR de 230, 300, 760, 860 y 2100 pb se observó en más de 63% de ellos; 90% de los individuos coamplificaron los fragmentos de IPCR de 300 y 860 pb (Figura 3).
DISCUSIóN
Inicialmente varios autores48,49 utilizaron la hibridación con la técnica Southern para determinar el tipo de integración del provirus HTLV-I; sin embargo, esta metodología además de ser dispendiosa y consumir tiempo tiene limitaciones de sensibilidad; por esta razón se desarrolló la IPCR con la gran ventaja de aumentar la sensibilidad en la determinación del número de provirus integrados. La IPCR es una herramienta que a partir de los estudios de Yoshida et al.2 en 1984 se ha utilizado como criterio diagnóstico para ATL desde 199443,44. Los resultados de la presente investigación sugieren que también se puede utilizar en PET/HAM.
Uno de los principales interrogantes en la biología de los retrovirus es su modalidad de integración al genoma de las células huéspedes. El HTLV-I es un retrovirus que principalmente integra su ADN proviral en células linfocitarias de tipo T. Sin embargo, existen estudios de integración proviral en otros tejidos como macrófagos, mucosa oral39-41 y componentes del sistema nervioso central42. En este trabajo se obtuvieron resultados solamente con respecto a la modalidad de integración proviral en PBMC. Es importante destacar que las diferencias encontradas en oligoclonalidad reflejan el proceso de integración en uno de los potenciales blancos celulares de la infección del virus in vivo. No se han efectuado hasta hoy estudios que determinen el patrón de clonalidad de provirus en el ADN de otros tipos celulares.
En este estudio se caracterizó la proliferación clonal de células infectadas por HTLV-I en todos los pacientes con PET/HAM, y se demostró que la modalidad de integración del HTLV-I en el genoma de individuos con PET/HAM es oligoclonal. Los resultados mediante IPCR confirman los obtenidos por Greenberg et al.32, Yoshida et al.2,3, Wattel et al.34 mediante Western Blot y los de Cavrois et al.35 entre otros autores38 que utilizaron IPCR. En todos ellos se ha definido una modalidad de integración oligoclonal del provirus HTLV-I en el genoma de pacientes PET/HAM.
Con respecto al número de amplicones obtenidos en este estudio se observó una tendencia en las personas con PET/HAM a poseer un mayor número de amplicones (7.2 ± 2.2) en comparación con las afectadas por otras enfermedades (4 ± 1) y con los portadores asintomáticos (4 ± 2). En parte, estas observaciones concuerdan con las evidencias ya informadas sobre las diferencias existentes en los patrones de clonalidad en la integración del HTLV-I entre individuos afectados con ATL donde se presenta de modo monoclonal y los que sufren PET/HAM, en quienes se ha informado un modo de integración oligoclonal2,3,27-29.
Otra de las características de la IPCR es que permite relacionar la intensidad de cada amplicón con su frecuencia para el genoma de cada individuo. Los resultados obtenidos en este trabajo sugieren que el retrovirus HTLV-I en los individuos del estudio, se integró en sitios específicos debido a la constancia en el patrón observado de bandas de integración. En este sentido son especialmente interesantes las bandas de 300 y 860 bp que se observaron en todos los pacientes con PET/HAM. Las observaciones de este artículo contrastan con el hecho generalmente aceptado de que el sitio de integración del HTLV-I es aleatorio2,3,32. Sin embargo, para confirmar es importante explorar las bandas a nivel de sus secuencias a fin de poder asegurar la existencia de sitios de integración preferencial del HTLV-I en personas con PET/HAM.
Las diferencias observadas en la intensidad de los ADN de IPCR en los individuos que presentaron integraciones múltiples, implicarían que si bien hay simultáneamente una expansión de varios clones portadores del provirus, algunos de ellos estarían más representados. Este hecho sugiere de nuevo la existencia de una preferencialidad en sitios de integración. Los presentes resultados aportan evidencias a las observaciones hechas por el grupo de Bernardi31 sobre la existencia de una preferencialidad parcial en el sitio de integración hacia los isocoros ricos en GC y se complementan con las observaciones de Chow et al.5 y de Wattel et al.34 quienes de acuerdo con sus estudios de clonación y secuenciación del ADN adyacente a los LTR de HTLV-I de células de pacientes con ATL mostraron que la región genómica flanqueante al provirus es por lo general rica en pares AT. En su conjunto los resultados sugirieron la existencia de sitios de integración preferencial del provirus HTLV-I en el genoma hospedero.
SUMMARY
The process of integration of HTLV-I in sixteen seropositive individuals grouped in eleven TSP/HAM patients, three asymptomatic carriers and two other pathology individuals was studied. The PBMC DNA of all individuals was submitted to IPCR and the results obtained showed evidences of integration and oligoclonal expansion patterns in infected HTLV-I individuals. Overall, the number of amplicons obtained by IPCR showed a tendency to exhibite a high number of integrations in TSP/HAM patients (7.2 ± 2) in comparison with asymptomatic carriers (4.0 ± 2.2) and individuals with other diseases included in the study (4.0 ± 1.0). Results obtained rendered evidence that TSP/HAM patient's infected cells have proliferated in oligoclonal manner. Taken together all results suggest a preferenciality of integration sites of provirus in PBMC of TSP/HAM patients.
REFERENCIAS
1. Poiesz BJ, Ruscetti FW, Gazdar AF, et al. Detection and isolation of type C retrovirus particles from fresh and cultured lymphocytes of a patient with cutaneous T-cell lymphoma. Proc Natl Acad Sci (USA) 1980; 77: 7415.
2. Yoshida M, Seiki M, Yamaguchi K, Takatsuki K. Monoclonal integration of human T-cell leukemia provirus in all primary tumors of adult T-cell leukemia suggests causative role of human T-cell leukemia virus in this disease. Proc Natl Acad Sci (USA) 1984; 81: 2534-2537.
3. Yoshida M. statement of the HTLV-I genome and its association with a unique T-cell malignancy. Biochim Byophis Acta 1987; 97: 145.
4. Gallo RC, Staal-Wong R. Retroviruses as etiologic agents of some animal and human leukemia and lymphomas as tools for elucidating the molecular mechanism of leukemogenesis. Blood 1982; 60: 545-557.
5. Chou KS, Okayama A, Su IJ, Lee TH, Essex M. Preferred nucleotide sequence at the integration target site of human T-cell leukemia virus type I from patients with adult T-cell leukemia. Int J Cancer 1996; 65: 20-24.
6. Zaninovic´ V, Biojó R, Barreto P. Paraparesia espástica del Pacífico. Colombia Med 1981; 12: 111-117.
7. Gessain A, Barin F, Vermant JC, et al. Antibodies to human T lymphotropic virus type I in patients with tropical spastic paraparesis. Lancet 1985; 2: 402.
8. Jacobson S, Mingoli ES, McFarlin D. Isolation of an HTLV-I like retrovirus from patients with tropical spastic paraparesis. Nature 1988; 331: 540-543.
9. Osame M, Usuku K, Izumo S, et al. HTLV-I associated myelopathy a new clinical entity. Lancet 1986; 1: 1031-1032.
10. Osame M, Igata A, Matsumoto M, et al. HTLV-I associated myelopathy (HAM). Kansenshogaku Zasshi 1988: 62S: 240-248.
11. Osame M, Matsumoto M, Usuku K, et al. Chronic progressive myelopathy associated with elevated antibodies to human T lymphotropic virus type I and adult T-cell leukemia-like cells. Ann Neurol 1987; 21: 117-122.
12. Mekhann G, Gibbs CJJr, Mora CA, et.al. Isolation and characterization of HTLV-I from symptomatic family members with tropical spastic paraparesis (HTLV-I encephalomyeloneuropathy). J Infect Dis 1989; 160: 371-379.
13. Rodger-Johnson P. Tropical spastic paraparesis/HTLV-I associated myelopathy. Etiology and clinical spectrum. Mol Neurobiol 1994; 8: 175-179.
14. Zaninovic´ V. El virus HTLV-I como posible causa de la paraparesia espástica tropical del Pacífico. Colombia Med 1986; 17: 2-8.
15. García-Vallejo F, Bonilla R, Gallego G, et al. Amplificación de segmentos de ADN del virus linfotrópico humano (HTLV-I) en pacientes con paraparesia espástica tropical y seropositivos asintomáticos. Colombia Med 1992; 23: 50-56.
16. Uchiyama T. Human T-cell leukemia virus type I (HTLV-I) and human diseases. Ann Rev Immunol 1997; 15: 15-37.
17. Trujillo CM, Concha M, Muñoz A, et al. Seroprevalence and cofactor of HTLV-I infection in Tumaco. AIDS Res Hum Retroviruses 1992; 8: 651-657.
18. García-Vallejo F, Soto L, Rengifo B, Chou K, Domínguez M, Essex M. Free human T-cell leukemia virus type I (HTLV-I) RNA templates in sera from patients with tropical spastic paraperesis/HTLV-I associated mielopathy is correlated with increased titers of IgM and IgA. J AIDS Human Retrovirol 1995; 10: 83.
19. García-Vallejo F. Molecular and immunological characteristics in the pathogenesis of tropical spastic paraparesis-HTLV-I associated myelopathy (TSP/HAM). Revista Ciencia Cultura 1996; 48: 351-358.
20. García-Vallejo F, Ramírez LS, Chou K, et al. Increased immunoglobulin levels in sera from patients with HTLV-I associated tropical spastic paraparesis in Tumaco, Colombia. AIDS Res Hum Retrovirol 1996; 10: 507.
21. García-Vallejo F, Balcázar N. Inmunogenicidad diferencial de dominios de las glicoproteínas de la envoltura del HTLV-I. Colombia Med 1996; 27: 101-105.
22. Gessain A, Gallo RC, Franchini G. Low degree of human T cell leukemia/lymphoma virus type I genetic drift in vivo as a means of monitoring viral transmission and movement of ancient human populations. J Virol 1992; 66: 2288-2295.
23. Kitze B, Usuku K, Izumo S, et al. Diversity of intrathecal antibody synthesis against HTLV-I and its relation to HTLV-I associated myelopathy. J Neurol 1996; 243: 393-400.
24. Bernardi G. The isochore organization of the human genome. Ann Rev Genet 1989; 23: 637-661.
25. Bernardi G. The isochore organization of the human genome and its evolutionary history. Ann Rev Genet 1995; 29: 445-476.
26. Yoshida M, Seiki M, Hattori S, Wattanabe T. Genome structure of human T-cell leukemia virus and its involvement in the development of adult T-cell leukemia. In Gallo RC, Essex ME, Gross L (eds.). Human T-cell leukemia/lymphoma virus. Cold Spring Harbor Laboratory, 1984. Pp. 141.
27. Wong-Stahl F, Hahn B, Manzari V, et al. A survey of human leukaemias for sequences of human retrovirus. Nature 1983; 302: 626-628.
28. Shinamoto Y, Miyahara M, Yamada H, et al. Adult T-cell leukaemia/lymphoma with multiple integrations of human T-cell lymphotropic virus type I proviral DNA: differing clinical features are linked to varied proviral integration. Br J Haematol 1996; 92: 632-638.
29. Shinamoto Y. Lack of correlation between T lymphotropic virus type I DNA integration and clinical course of adult T-cell leukemia/lymphoma. Blood 1995; 85: 1673-1674.
30. Seiki M, Eddy R, Shows TB, Yoshida M. Non-specific integration of HTLV provirus genome into adult T-cell leukemia cells. Nature 1984; 309: 640-642.
31. Glukhova LA, Zoubak SV, Rynditch AV, et al. Localization of HTLV-I and HIV-I proviral sequences in chromosomes of persistently infected cells. Chromosome Res 1999; 7: 177-183.
32. Greenberg SJ, Jacobson S, Waldman TA, et al. Molecular analysis of HTLV-I proviral integration and T-cell receptor arrangement indicates that T-cells in tropical spastic paraparesis are polyclonal. J Infect Dis 1989; 159: 741-743.
33. Yoshida M, Osame M, Kawai H, et al. Increased replication of HTLV-I in HTLV-I associated myelopathy. Ann Neurol 1989; 26: 331.
34. Wattel E, Vartarian JP, Wain-Hobson S. Clonal expansion of HTLV-I infected cells in asymptomatic and symptomatic carriers without malignancy. J Virol 1995; 69: 2863-2868.
35. Cavrois M, Leclercq I, Gout O, Gessain A, Wain-Hobson S, Wattel E. Persistent oligoclonal expansion of human T-cell leukemia virus type 1-infected circulating cells in patients with tropical spastic paraparesis/HTLV-1 associated myelopathy. Oncogene 1998; 17: 77-82.
36. Sambrook J, Fritzch EF, Maniatis T. Molecular cloning. A laboratory manual. Cold Spring Harbor Press, 1989.
37. Takemoto S, Matsuoka M, Yamaguchi K, et al. A novel diagnostic method of adult T-cell leukemia: monoclonal integration of human T-cell lymphotropic virus type I provirus DNA detected by inverse polymerase chain reaction. Blood 1994; 4: 3080-3085.
38. Cerón-Souza I, Barreto G, García-Vallejo F. Patrones de integración del virus linfotrópico humano tipo I (HTLV-I) en células linfocitarias de individuos seropositivos. En: Resúmenes del XXXIII Congreso Nacional de Ciencias Biológicas, Ibagué, 1998.
39. Soto-Ramírez LE, García-Vallejo F, Vergara A, Borrero I, Marlink R, Essex M. Antibodies to HTLV-I in gingival crevicular fluid of TSP and seropositive individuals from Tumaco, Colombia. AIDS Res Hum Retrovirus 1994; 10: 446.
40. Soto-Ramírez LE, García-Vallejo F, Vergara A. Antibodies to HTLV-I gingival crevicular fluid of TSP and seropositive individuals from Tumaco, Colombia. Viral Immunol 1995; 8: 141-150.
41. González N, Blank A, Zaninovic´ V, García-Vallejo F. Respuesta inmune de mucosa oral en pacientes PET/MAH. En: Resúmenes del XXXIII Congreso Nacional de Ciencias Biológicas, Pamplona 1997.
42. Kira JL, Koyonagi Y, Yamada T, et al. Increased HTLV-I proviral DNA in HTLV-I associated myelopathy: A quantitative polymerase chain reaction study. Ann Neurol 1991; 29: 194-201.
43. Furukawa J, Fujisawa J, Osame M, et al. Frequent clonal proliferation of human T cell leukemia virus type I (HTLV-I) infected T-cells in HTLV-I/associated myelopathy (HAM-TSP). Blood 1995; 80: 1012-1016.
44. Yamaguchi K. Polyclonal integration of HTLV-I proviral DNA in lymphocytes from seropositive individuals: an intermediate state between the healthy carrier state and smouldering ATL. Br J Haemathol 1988; 68: 169-174.
45. Renjifo B, Soto L, García-Vallejo F, Essex M. Human T-cell leukemia virus type I (HTLV-I) molecular genotypes and disease outcome. J AIDS Hum Retrovirol 1996; 13S: 146-152.
46. Yoshida M, Osame M, Usuku K, et al. Virus detected in HTLV-I associated myelopathy and adult T-cell leukemia are identical on DNA blotting. Lancet 1987; 1: 1085.
47. Yamaguchi K, Seiki M, Yoshida M, et al. The detection of human T-cell leukemia virus proviral DNA and its application for classification and diagnosis of T-cell malignancy. Blood 1984; 63: 235.
48. Triglia T, Peterson MG, Kemp DJ. A procedure for in vitro amplification of DNA segments that lie outside the boundaries of known sequences. Nucleic Acids Res 1988; 16: 8186.
49. Silver J. Inverse polymerase chain reaction. In McPherson MJ, Quirke P, Taylor GR (eds.). PCR: A practical approach. New York, Oxford: Academic Press, 1991. Pp. 137.