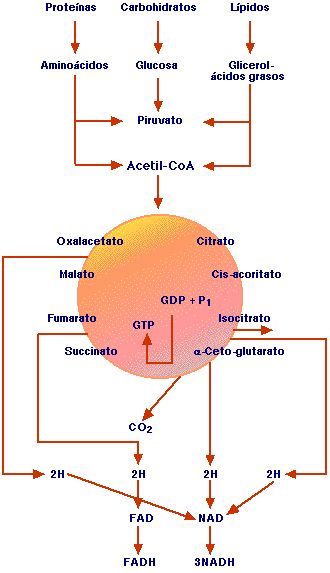
Figura 1. Ciclo de Krebs. Producción de acetil CoA, a partir de los diversos sustratos energéticos, oxidación del acetil CoA en el ciclo de Krebs, con producción de dos moléculas de CO2, 3 NADH y 1 FADH y un GTP.
Carlos Alberto Ordoñez, M.D.
Docente Adjunto, Departamento de Cirugía, Facultad de Salud, Universidad del Valle, Cali, Colombia
El adenosín trifosfato (ATP), es la moneda energética de los seres vivos. Para poder ser sintetizado, los organismos requieren oxidar los sustratos energéticos de la dieta, proteínas, grasas y carbohidratos. Inicialmente estas sustancias tienen vías metabólicas separadas hasta alcanzar en su degradación un metabolito común que es el acetil CoA. A partir de este punto entran al ciclo de Krebs, con producción de CO2 e hidrogeniones, estos últimos se transportan por óxido reducción a la cadena respiratoria donde se formará agua endógena y ATP. Para lograr esta oxidación de los sustratos con alta producción de energía, es indispensable el oxígeno que actúa como comburente en las reacciones.
The adenosine triphosphatase [5 (pyro-) triphosphate of adenosine], ATP, is like the energy currency among living beings. For its synthesis, all the organisms need to oxydize energetic substrata of diet, proteins, lipids and carbohydrates. Initially these substances have separate metabolic ways until during their degradation they reach a common metabolite, acetyl coenzyme A, CoA. From this point on, they enter to Krebs' cycle, producing CO2 and hydrogen ions that will be transported by oxidation-reduction to the respiratory chain where endogen water and ATP will be formed. For obtaining this substratum oxidation, with a high energy production, it is necessary the presence of oxygen acting as a burner at the reactions.
La energía adquirida por las células se conserva en ellas para ser utilizada principalmente cuando se requiera en forma de adenosín trifosfato (ATP). Tanto si proviene de la luz solar o de la oxidación de compuestos orgánicos, se invierte en la formación de ATP, en una proporción muy alta. El ATP es entonces el "fluido energético" que pondrá en marcha las demás funciones de la célula.
Todos los seres vivos necesitan un aporte continuo de materia y energía, aunque existen grandes diferencias en la forma de obtenerlas y de su utilización. Los vegetales son seres autótrofos, utilizan la energía solar como fuente de energía y como materia usan el agua, el dióxido de carbono (CO2) y los iones orgánicos. En la fotosíntesis los cloroplastos captan la energía solar y tienen la maquinaria para convertirla en energía química (ATP); además extraen los iones de hidrógeno del agua para convertirlos en equivalentes de reducción (NADPH + H). Como producto de esta reacción se libera oxígeno molecular.
Con el ATP, los NAPDH + H y el CO2 en un proceso cíclico forman la glucosa y a partir de esta, con fuentes inorgánicas de nitrógeno, los vegetales tiene la capacidad de sintetizar todas las biomoléculas orgánicas (azúcares, lípidos, proteínas y ácidos nucleicos).
El hombre y los animales, seres heterótrofos utilizan como fuente de energía y materia las biomoléculas sintetizadas por los vegetales, ingeridas en los alimentos directamente. Las biomoléculas ingeridas por el hombre se degradan metabólicamente hasta convertirse en CO2 y H2O, y derivados nitrogenados, que liberan energía química (ATP). Esta energía se utiliza para la realización de trabajo y la síntesis proteica.
La materia sufre una serie de transformaciones cíclicas, pues pasa de los vegetales al hombre y de estos a través del suelo y la atmósfera de nuevo a los primeros. El flujo de energía solar es unidireccional: se inicia como energia solar y se degrada como energía térmica (calor) pero sin destruirse1.
El conjunto de intercambios y transformaciones de materia y energía que tiene lugar en el ser vivo recibe el nombre de metabolismo. La oxidación de las moléculas complejas, con eliminación de productos de desecho y liberación de energía, se llama catabolismo; y la biosíntesis de sustancia propia a partir de moléculas sencillas, con gasto de energía, se denomina anabolismo.
Los alimentos ingeridos en la dieta son macromoléculas de almidón, proteínas y triglicéridos que en la digestión se hidrolizan a monómeros, como monosacáridos, aminoácidos, ácidos grasos y glicerol. Estos monómeros en las células se absorben y se incorporan o entran para ser oxidados con producción de energía o se derivan a la biosíntesis de nuevo material celular con consumo de energía.
Una vez que se absorbe la glucosa en el hígado, se fosforila y se almacena en forma de glucógeno o se metaboliza y entra en la vía glucolítica, para convertirse en dos moléculas de piruvato y éste en lactato sin que haya necesidad de oxígeno. Pero, en condiciones aeróbicas el piruvato se convierte en acetil CoA que entra al ciclo de Krebs (Figura 1).
Figura 1. Ciclo de Krebs. Producción de acetil CoA, a partir de los diversos sustratos energéticos, oxidación del acetil CoA en el ciclo de Krebs, con producción de dos moléculas de CO2, 3 NADH y 1 FADH y un GTP.
Los destinos de degradación de los aminoácidos son la oxidación a CO2, gluconeogénesis y cetogénesis. Luego que se extrae el grupo amino por medio de la transaminación y la desaminación, se da origen a piruvato, acetil CoA y a otros diversos intermediarios en el ciclo de Krebs.
El proceso general por cuyo medio se oxidan los ácidos grasos se llama -oxidación, pues pierden fragmentos de 2 carbonos (acetil CoA), empezando por el carbono b y se reduce el ácido graso hasta el final de la cadena, para quedar todo convertido en acetil CoA a fin de entrar al ciclo de los ácidos tricarboxílicos.
El acetil CoA es un metabolito intermediario de gran importancia, pues es el punto de confluencia de la oxidación de los azúcares, lípidos y proteínas. El acetil CoA se oxida por completo a CO2 y H2O en presencia de O2, por medio de un proceso enzimático cíclico conocido como ciclo de Krebs, o ciclo de los ácidos tricarboxílicos o ciclo del ácido cítrico2.
El ciclo de Krebs tiene lugar en las mitocondrias de las plantas y animales, mientras que en los procariotas ese ciclo ocurre en el citosol; se hace, pues en todos los organismos aerobios. El punto de entrada de todos los combustibles al ciclo de Krebs a través del intermediario metabólico acetil CoA; éste se condensa con una molécula de oxalacetato, para dar citrato, de allí el nombre de ciclo del ácido cítrico, y como da origen a otros ácidos tricarboxílicos, también se llama ciclo de los ácidos tricarboxílicos. El oxalacetato que se desprende en forma de CO2 corresponden al oxalacetato y no al último acetilo incorporado.
En consecuencia, el oxalacetato que se regenera al final del ciclo no contiene los mismos átomos de carbón que el oxalacetato original. En cada vuelta del ciclo se oxida un residuo acetil CoA a dos moléculas de CO2; simultáneamente se reducen 4 coenzimas 3 NAD+ a 3 NADH y 1 FAD a 1 FADH2. Además se genera un GTP a partir de GDP + fosfato inorgánico que dará posteriormente un ATP3.
La velocidad de las enzimas para regular el ciclo depende básicamente de la cantidad de ATP; si hay demasiado, la velocidad de ciclo disminuye y, si por el contrario hay exceso de ADP la velocidad del ciclo aumenta4. El ciclo de Krebs representa la vía final común de la oxidación aeróbica de todos los sustratos de la dieta (proteínas, lípidos y carbohidratos) con producción de CO2 como desecho, reducción de coenzimas que van a transportar átomos de hidrógeno y electrones que se utilizarán en la cadena respiratoria para la formación de ATP y de una molécula de GTP que reaccionará con el ADP y formará ATP. Así se resume la utilidad y la productividad del ciclo.
Cada molécula de NAD+ acepta 2 electrones y 1 protón. El protón y uno de los electrones se une a un átomo de carbono de la molécula de NAD+; el otro electrón neutraliza la carga positiva. Esta forma reducida de NAD+ se denomina NADH. El NADH es el principal intermediario entre el ciclo de Krebs y la cadena respiratoria.
Las enzimas de la membrana interna de la mitocondria que transportan hidrógeno y electrones componen la cadena respiratoria y este conjunto de pasos hasta llevar los electrones al O2 y formar agua, se llama respiración interna5,6. La teoría quimiosmótica7,8 sobre la respiración mitocondrial, que es la más aceptada, explica la formación de ATP de la siguiente manera (Figura 2).
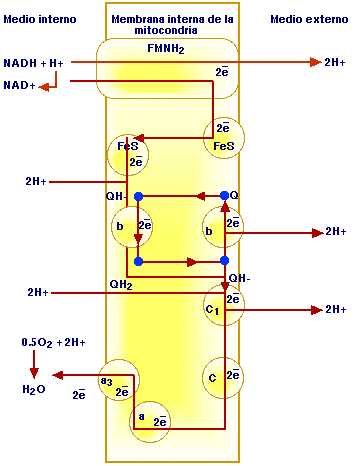
Figura 2. Cadena respiratoria. Parte 1.
En la membrana de la mitocondria el NADH cede 2 electrones y 1 protón a un grupo transferidor denominado flavín-mononucleótido (FMN). En este proceso se oxida el NADH, es decir, retorna a la forma de NAD+ y el FMN al haber aceptado 2 electrones y 1 protón, capta un protón adicional del medio interno con lo que se reduce a FMNH2 que tiene completos 2 átomos de hidrógeno. La molécula de FMN está unida a una proteína de gran tamaño que atraviesa por entero la membrana. El FMNH2 transfiere los 2 átomos de hidrógeno desde el interior de la membrana al exterior. Los átomos se ionizan y los protones se liberan al medio extramitocondrial, con lo que se tienen los 2 primeros protones (2H+) liberados. Los 2 electrones se transfieren a la superficie interna de la membrana a través de una ferroproteína sulfurada (FeS). El FMNH2 al ceder 2 protones y 2 electrones retorna a su forma original y puede de nuevo ser reducido por el NADH.
Las ferroproteínas ceden los electrones a 2 moléculas de ubiquinona (Q), o coenzima Q; cada una de ellas adquiere un protón del medio interno y da lugar a la forma semiquinona (QH*). La semiquinona capta 2 electrones más, suministrados por el citocromo ß y con 2 protones más procedentes del medio interno de la mitocondria, da lugar a la forma hidroquinona (QH2) que es el estado más reducido. Cada hidroxiquinona cede un electrón al citocromo C1 que es la siguiente proteína de la cadena respiratoria y libera un protón más, siendo hasta el momento 4 los protones liberados (4H+). Todas las moléculas de ubiquinona se encuentra en el estado de semiquinona (QH*). Completan el ciclo y retorna al estado de máxima oxidación (Q). Cada una cede un protón restante al medio externo y transfiere al citocromo ß el electrón asociado. Así se liberan los otros 2 protones, y se completan los 6 que se deben expulsar de la mitocondria a través de la membrana. Los 2 electrones restantes son devueltos al ciclo por medio del citocromo C1 atraviesan los citocromos C, A, A3, y por último el A3 es oxidado por oxígeno molecular. Los 2 electrones son cedidos a un átomo de oxígeno y 2 protones son captados del medio interno de la mitocondria con lo que se forma una molécula de agua (H2O endógena).
Durante esta larga serie de reacciones de óxido reducción, el par de electrones atraviesa 3 veces la membrana en ambos sentidos y entre 2 protones en cada una de las salidas, en total 6 (6H+), con formación de una molécula de agua al suministrar los 2 electrones al oxígeno.
Así se ha producido un gradiente de protones a través de la membrana mitocondrial. Debido a este gradiente, los protones del exterior experimentan una cierta tendencia a volver al interior del compartimiento. Como la membrana es impermeable a los protones, su entrada se produce mediante la ATP-sintetasa que es una proteína transportadora de H+. Los protones adquirieron un potencial energético al ser transportados fuera de la membrana, en contra del gradiente (Figura 3).
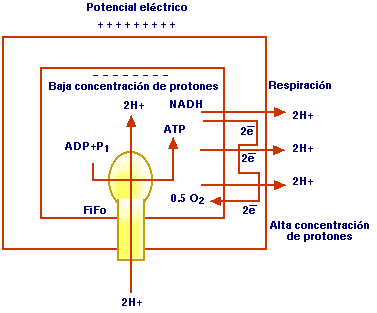
Figura 3. Cadena respiratoria. Parte 2. Fosforilación oxidativa. FiFo = ATP sintetasa
El flujo de protones a favor del gradiente es un proceso exoergónico y la energía liberada se utiliza en parte para la fosforilación del adenosín difosfórico (ADP), y así se forma el ATP. Se genera un ATP por cada 2H+ que atraviesan la membrana; la reoxidación de un NADH produce 3 ATP, mientras que 1 FADH, origina 2 ATP5-8.
El equilibrio energético de las vías oxidativas se puede ejemplificar si se mira la producción de ATP al oxidar una molécula de glucosa que es de 36 ATP. Por ejemplo, el ácido palmítico en su oxidación completa produce 129 ATP. El glutamato, que es el receptor final de casi todos los grupos amino de los aminoácidos, tiene un total de 27 ATP.
Esto permite evaluar el rendimiento energético en la degradación de las diferentes biomoléculas4. El nucleótido adenosín trifosfato (ATP) es el compuesto principal que almacena y transporta la energía libre y lo hace a través de la creación y ruptura de enlaces ricos en energía. Se llama energía de enlace a la energía libre de Gibbs que se desprende al hidrolizar una molécula de ácido fosfórico del ATP1.
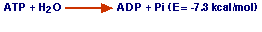
El ATP es el producto final de la conservación y la transferencia de energía en el metabolismo oxidativo de todos los sustratos y al hidrolizarse liberará la energía almacenada en los procesos anabólicos del organismo. Así se produce el equilibrio entre la producción y el consumo de energía para mantener la vida.
1) El ATP es, excepto:
a. Adenosín trifosfato.
b. El fluido energético
c. Energía química.
d. Transporta energía libre.
e. ATP + Pi ------>H2O + ADP
2) Lo siguiente es cierto sobre metabolismo, excepto:
a. En la fotosíntesis la energía solar se convierte en energía química.
b. Los cloroplastos con ATP, NADPH, H+, CO2, forman glucosa.
c. La energía tiene un flujo unidireccional y se destruye.
d. Los intercambios de materia y energía en el ser vivo se llaman metabolismo.
e. La oxidación de las moléculas complejas, con liberación de energía se llama catabolismo.
3) Todo es cierto en metabolismo, excepto:
a. Los alimentos de la dieta son macromoléculas de almidón, proteínas y triglicéridos.
b. La glucosa se convierte en piruvato y lactato en anaerobiosis.
c. Hay aminoácidos glucogénicos.
d. La ß-oxidación convierte los ácidos grasos en acetil CoA.
e. El lactato entra directo al ciclo de Krebs.
4) Todo es cierto con respeto al ciclo de Krebs, excepto:
a. Es la vía final común de la oxidación aeróbica de todos los sustratos de la dieta.
b. La velocidad de las enzimas para regular el ciclo depende de la cantidad de ATP.
c. Se realiza en el citosol de las plantas y animales.
d. El ciclo produce 2CO2, 3NADH, GTP.
e. Se llama también ciclo de ácido cítrico.
5) Todo es cierto en la cadena respiratoria, excepto:
a. Es una cadena que transporta electrones y protones.
b. El NADH es el principal intermediario entre el ciclo de Krebs y la cadena respiratoria.
c. El último en aceptar electrones es el oxígeno y así se forma el agua.
d. El ATP no se forma en la cadena respiratoria sino en el ciclo de Krebs.
e. La fosforilación oxidativa es la cadena respiratoria.
6) Todo es cierto respecto al ciclo de la materia y la energía, excepto:
a. En las reacciones luminosas de la fotosíntesis, los cloroplastos arrancan los electrones e hidrogeniones del agua y con la energía solar forma ATP.
b. Los seres heterótrofos producen CO2, H2O, amoníaco, y los seres autótrofos los utilizan para formar compuestos orgánicos y O2.
c. El hombre sintetiza ATP en la oxidación anabólica de los compuestos orgánicos.
d. El ATP se utiliza para realizar trabajo mecánico, biosíntesis de proteínas, transporte activo.
e. La energía no se crea ni se destruye y el ser vivo tiende a la entropía.
7) Todo es cierto respecto al metabolismo, excepto:
a. Los aminoácidos producen piruvato, acetil CoA, NH3.
b. Los lípidos de la dieta producen ácidos grasos y glicerol.
c. La glicólisis aeróbica da 2 moléculas de lactato antes del ciclo de Krebs
d. El ciclo de los ácidos tricarboxílicos tiene como metabolito intermediario al acetil CoA
e. El oxalacetato cede 2 carbonos en cada vuelta del ciclo.
8) Todo es cierto para el ciclo de Krebs y la cadena respiratoria, excepto:
a. Cada vuelta del ciclo produce 2CO3, 3NADH, 1 FADH2, 36 ATP.
b. El ciclo reduce las coenzimas que van a transportar protones.
c. Las enzimas de la membrana interna de la mitocondria que transportan hidrogeniones y electrones forman la cadena respiratoria
d. Los electrones (2) son suministrados al O2 para formar H2O.
e. Los citocromos hacen parte de las reacciones de óxido reducción.
9) Todo es cierto en la producción de energía, excepto:
a. La ATP-sintetasa es una proteína transportadora de H+.
b. El flujo de protones es un proceso exoergónico.
c. Se genera un ATP por cada 2 H+ que atraviesan la ATP-sintetasa.
d. Al oxidar una molécula de glucosa se producen 36 ATP.
e. La cadena respiratoria es un proceso endergónico.
10) Todas las afirmaciones son ciertas, excepto:
a. La energía libre de Gibbs es la que se desprende al hidrolizar una molécula de ATP.
b. ATP + H2O ------>ADP + Pi (E= -7.3 kcal/mol).
c. El ATP es el producto final de conservación y transferencia de energía en el metabolismo oxidativo.
d. Sin O2suficiente las células no podrán sintetizar el ATP requerido para tamponar los hidrogeniones.
e. El ATP es la única molécula que almacena energía.
1.e 2.c 3.e 4.c 5.d 6.c 7.c 8.a 9.e 10.e
1. Macarulla JM, Goñi FM. Introducción al metabolismo. Bioquímica humana. Barcelona; Editorial Reverte, 1987. Pp. 197-218.
2. Felig P, Havel R, Smith L. Metabolismo. En Fisiopatología de Smith/Their. 2ª ed. Buenos Aires; Editorial Médica Panamericana, 1990. Pp. 303-27.
3. Hanoune J. Las regulaciones metabólicas. En Fisiología humana. Meyer P (ed). Barcelona; Editorial Salvat, 1985. Pp. 59-89.
4. Sánchez PJ. Equilibrio ácido básico. Conferencia Unidad de Cuidados Intensivos. Hospital San Juan de Dios, Bogotá.
5. Macarulla JM, Goñi FM. Vías metabólicas degradativas. Fosforilación oxidativa. En Bioquímica humana. Barcelona; Editorial Reverte, 1987. Pp. 245-62.
6. Mayes P. Fosforilación oxidativa y sistema de transporte mitocondrial. En Bioquímica de Harper. Editorial El Manual Moderno, 1984. Pp. 154-61.
7. Guyton AC. Metabolismo de los carbohidratos y formación de triofosfato de adenosina. En Fisiología médica de Guyton, 6ª ed. México; Editorial Interamericana, 1984. Pp. 993-1004.
8. Hinkle P, McCarty R. Cómo fabrican ATP las células. Sci Am 1978; 11: 58-75.