Perfil fisiológico de la leptina
Julio César Sánchez, M.D., M.Sc., Ph.D.*
* Profesor Asociado, Departamento de Ciencias Básicas, Facultad de Salud, Universidad Tecnológica de Pereira.
e-mail: jcsanchez@utp.edu.co Recibido para publicación junio 7, 2004 Aprobado para publicación enero 28, 2005
RESUMEN
INTRODUCCION: La leptina fue descubierta en el contexto de sus efectos sobre el control de la alimentación y del gasto energético. Aunque en un principio se pensó que la leptina actuaba sólo sobre ciertos centros hipotalámicos, en la actualidad le ha sido asignada una mayor diversidad de funciones.
MOLÉCULA Y RECEPTOR: La leptina es una proteína de la familia de las citocinas y su receptor, similar al de éstas. Aunque existen al menos seis isoformas del receptor, la forma activa corresponde al subtipo ObRb, asociado fundamentalmente con el sistema JAK2-STAT3 y con otras cascadas de señalización intracelular.
FUNCIONES: La leptina participa en procesos tales como regulación del peso corporal, de la alimentación y del gasto energético, reproducción, crecimiento, función inmune, tono vascular y probablemente muchos otros aún por determinar.
ENFERMEDADES ASOCIADAS: La leptina se ha asociado con un amplio número de procesos patológicos que incluyen cáncer de diversos tipos, así como también enfermedades psiquiátricas, hipertensión arterial, diabetes mellitus no insulinodependiente, transtornos reproductivos, insuficiencia placentaria, inmunodeficiencias y osteoartritis.
CONCLUSIONES: La leptina es un regulador hormonal de muchos procesos biológicos y como tal interviene en la fisiopatología de un amplio número de enfermedades.
Palabras clave: Receptor de leptina; Regulación hormonal.
SUMMARY
INTRODUCTION: Leptin was discovered because of its actions in the control of feeding and metabolic expenditure, affecting hypothalamic nucleus. Since that, a wide diversity of additional functions have been described for this hormone.
MOLECULE AND RECEPTOR: Leptin is a protein that belongs to the cytocine family and hence its receptor is similar to cytocine receptors. There are at least six isoforms, although ObRb subtype is the active form. ObRb is associated with JAK2-STAT3 system and also activates other intracellular signalling systems.
FUNCTIONS OF LEPTIN: Leptin is involved in a wide variety of processes such as regulation of body weight, regulation of feeding and metabolic expenditure, reproduction, growth, immune function, vascular tone and probably in many others to be determined.
ASSOCIATED DISEASES: Leptin has been associated with a number of diseases including cancer of various types, psichiatric illnesses, arterial hypertension, non-insulin-dependent diabetes mellitus, reproductive alterations, placenta insufficiency, immune defficiencies and osteoarthritis.
CONCLUSIONS: Leptin is an hormonal regulator of many biological processes and can be involved in the pathophysiology of a wide number of diseases.
Key words: Leptin receptor; Hormonal regulation.
Los informes acerca de la existencia de un sistema fisiológico regulador del peso corporal, de la alimentación y del gasto de energía se iniciaron desde hace aproximadamente cuatro décadas. En la mayoría de los mamíferos, la masa de tejido adiposo tiende a permanecer constante durante largo tiempo. Esto hizo sospechar la existencia de algún mecanismo homeostático encargado por una parte, de vigilar las variaciones en el gasto energético y por otra, de generar cambios compensatorios en la ingesta de alimentos, además de balancear las reservas energéticas en forma de tejido adiposo1. Esta afirmación es consistente con observaciones experimentales y clínicas que demuestran que, después de una disminución importante de la adiposidad secundaria a dietas hipocalóricas extremas o a remoción quirúrgica, el peso corporal se restaura en un período relativamente corto si se permite un acceso libre a la comida2. El fenómeno inverso también ocurre, de modo que cuando un individuo sometido a alimentación forzada aumenta de peso, una vez cesa la intervención, su apetito disminuye y por ende, también su ingesta de alimentos, hasta recuperar el peso original2.
Las observaciones clásicas hechas por Hetterington y Ransom3y Hervey4sugirieron que el sistema regulador propuesto estaba relacionado con el hipotálamo, porque ratas con lesiones en el área ventromedial desarrollaban apetitos exagerados y obesidad mórbida. Además, las ratas normales sometidas a circulación cruzada con los animales sometidos a dichas lesiones perdían completamente el apetito y finalmente murieron de inanición, no obstante tenían acceso al alimento. Estos hechos demostraron la existencia de un factor circulante que producía ausencia del apetito en las ratas normales y que además, estaba aumentado en las ratas obesas, a consecuencia del daño hipotalámico.
Otras investigaciones en torno al concepto de �factor de saciedad circulante� condujeron al descubrimiento en el ratón de dos mutaciones denominadas ob (obese) y db (diabetes), responsables de producir alteraciones tales como hiperfagia, hiperinsulinemia, hiperglicemia, gasto metabólico reducido, hipogonadismo, infertilidad y obesidad de inicio temprano conducente a obesidad mórbida5. Cuando los ratones ob/ob eran sometidos a circulación cruzada con ratones normales, no ganaban peso. Sin embargo, al efectuar el mismo procedimiento entre ratones db/db y ratones normales, los últimos desarrollaron una severa hipofagia6. Estas observaciones permitieron a Coleman6concluir en la década de 1970 que los ratones ob/ob no producían un �factor de saciedad humoral� y que los ratones db/db no sintetizaban el producto adicional requerido para la respuesta a dicho factor. Mientras que a los genes responsables de las mutaciones mencionadas se les llamó también ob y db, al producto del gen ob se le denominó leptina (del griego leptos o delgado). Por otra parte, el producto del gen db es el mismo receptor de la leptina, indispensable para su acción a nivel celular. Con posterioridad se logró caracterizar a la leptina y clonar los genes db y ob en ratones y en seres humanos7. Sin embargo, la concepción inicial restringida de la leptina como hormona antiobesidad, fue reemplazada por una más compleja que la caracteriza como regulador neuroendocrino de una gran diversidad de procesos.
LA MOLÉCULA DE LA LEPTINA, SU RECEPTOR Y LA SEÑALIZACIÓN INTRACELULAR ASOCIADA
La leptina es un péptido glucosilado de 16 KD. Está constituido por 146 aminoácidos y contiene un puente disulfuro necesario para su actividad biológica. Posee un alto grado de homología entre especies (84% entre la leptina humana y murina, por ejemplo)7, además de compartir algunas similitudes estructurales con las citocinas. Aunque es producida en respuesta a una diversidad de estímulos, no se ha demostrado la existencia de depósitos intracelulares de esta hormona, lo que sugiere que los estímulos actuarían sobre la síntesis, mas no sobre la secreción de la molécula.
El gen ob está localizado en el cromosoma7del ser humano y en el cromosoma 6 del ratón7. Cuando se producen mutaciones en ob, los animales de experimentación desarrollan el fenotipo típico ob/ob. El gen posee 650 kilobases, 3 exones y 2 intrones, mientras que su ARNm posee 4.5 kilobases. Las mutaciones de ob son muy raras en los seres humanos8. Los individuos afectados desarrollan un cuadro de hiperfagia, obesidad mórbida e hipogonadismo hipotalámico pero, a diferencia de los ratones ob/ob, no desarrollan hiperinsulinemia o hiperglicemia. Las razones para estas diferencias fenotípicas son desconocidas hasta el momento.
El producto del gen db, el receptor de la leptina (ObR o LR), es expresado en todos los tejidos estudiados con miras a su identificación9, lo que indica la importancia de la leptina como hormona reguladora en todos los tipos celulares.
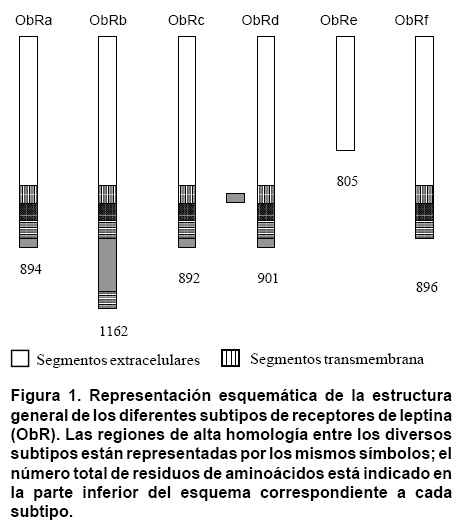
Las mutaciones del gen db son también muy raras en los seres humanos. Clement et al.10 describieron algunos casos en los que miembros de una misma familia presentaban hiperfagia, obesidad mórbida e hipogonadismo hipotalámico, sin hiperinsulinemia acompañante. Entre las seis isoformas conocidas del receptor se encuentran ObRa-f, producto de múltiples combinaciones de diferentes segmentos del gen11 y ObR, perteneciente a la familia de receptores de las citocinas clase 1, que también incluye a los receptores de la interleucina 6 y al factor estimulante del crecimiento de colonias (GCSF). Todas las isoformas del receptor comparten un dominio extracelular idéntico, localizado en el extremo aminoterminal, donde se encuentra el sitio de unión con la leptina. Sin embargo, ellas difieren en su porción carboxiterminal (Figura 1). Cinco de las seis isoformas (a, b, c, d y f) poseen dominios transmembranales, pero sólo ObRb contiene el segmento intracelular necesario para la activación de las cinasas12.
Las mutaciones responsables del fenotipo db/db determinan la incapacidad para sintetizar ObRb. La isoforma ObRe sólo posee el segmento extracelular y no está asociada con las membranas, pues se encuentra en forma circulante11. ObRb es conocida como la forma �larga� o �completa� del receptor, mientras que las demás isoformas corresponden a las formas �cortas� del mismo.
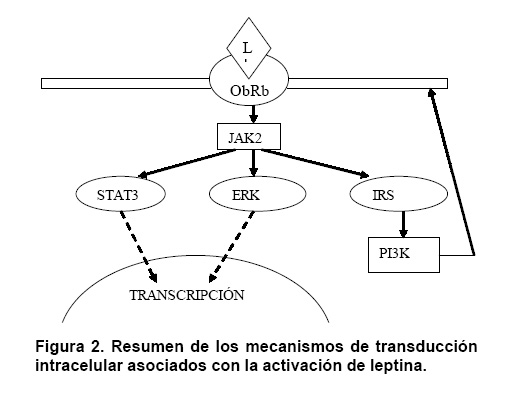
ObR existe como un homodímero preformado. Como ocurre con todos los receptores tipo citocina, la unión del ligando con ObR da lugar a la activación de señales intracelulares, aunque el receptor per se no posee actividad enzimática intrínseca. La señalización intracelular mediada por ObRb está asociada con el sistema JAK-STAT12. Las JAK (del inglés Janus Kinases) son una familia de tirosíncinasas intracelulares con dos sitios activos, característica que evoca a Janus, el dios griego de las dos caras. Las JAK son activadas principalmente por receptores tipo citocina. Hasta la fecha han sido identificados cuatro miembros de esta familia: Tyk2, JAK1, JAK2 y JAK3. Los tres primeros están ampliamente distribuídos, mientras JAK3 sólo es expresado en células inmunitarias y hematopoyéticas. ObRb activa de manera preferencial a JAK213, lo que induce autofosforilación del complejo LRb-JAK2. Este evento da inicio a una cascada de fosforilación que involucra a diversos sistemas enzimáticos. Como todas las tirosíncinasas, JAK2 fosforila a las proteínas con dominios SH2. Estas proteínas pertenecen a tres familias diferentes. La primera familia está constituída por las proteínas ERK (Extracellular-signal Regulated Kinase). La segunda familia por su parte, comprende las proteínas STAT3 (Signal Transducers and Activators of Transcription 3) las que, una vez fosforiladas, son translocadas al núcleo donde funcionan como reguladoras transcripcionales. El tercer grupo de proteínas asociado al sistema LR-JAK2 incluye a las IRS (Insulin Receptor Substrates), pertenecientes a su vez al grupo de proteínas muelle (docking proteins). Las IRS son fosforiladas por varios sistemas tirosina kinasa, entre los que se incluyen los asociados a los receptores de insulina y algunos receptores de citocinas. Las proteínas IRS, aunque no poseen actividad enzimática, pueden ser fosforiladas en múltiples residuos de tirosina. Su nombre se debe a que median en el reclutamiento o anclaje de las proteínas SH2, hecho que a su vez, conduce a la activación de cascadas de fosforilación. Las proteínas IRS están relacionadas con la activación de la PI 3 cinasa (fosfatidilinositol 3 cinasa). Esta enzima actúa sobre los fosfolípidos de la membrana plasmática, con la finalidad de permitir la síntesis de IP3, conocido mediador de la liberación de calcio a partir de los depósitos intracelulares (Figura 2). La activación de ObRb también puede conducir a la estimulación de la vía de señalización Ras-MAPK en varios tipos celulares14, vía que parece revestir una menor importancia.
Mediante estudios electrofisiológicos ha sido demostrado que la leptina también tiene efectos inmediatos sobre las neuronas hipotalámicas15 y sobre las células b del páncreas16. Este efecto depende, tanto de la activación del receptor, como de la de los canales de potasio sensibles a ATP (KATP) y, en apariencia, no guarda relación alguna con la activación de la transcripción. Por otra parte, la leptina podría activar canales de potasio dependientes de calcio en neuronas del hipocampo17.
REGULACIÓN DE LA SÍNTESIS Y LA LIBERACIÓN DE LA LEPTINA
La leptina es producida principalmente en los adipocitos18, aunque también ha sido hallada en otros órganos y tejidos, tales como hipotálamo, hipófisis, placenta, músculo esquelético, mucosa gástrica y epitelio mamario19. La regulación de la expresión de la proteína depende de manera importante de los depósitos grasos del organismo. Así, los adipocitos de mayor tamaño producen más leptina, mientras que los adipocitos epiploicos secretan menos que los subcutáneos. La cantidad de triglicéridos almacenados en el adipocito es también proporcional a la cantidad de leptina producida por cada adipocito. Por esta razón, los niveles circulantes de leptina son proporcionales a la cantidad de grasa corporal.
La secreción de leptina varía de acuerdo al ritmo circadiano. La proteína es secretada en forma de pulsos, con una frecuencia aproximada de un pulso cada 45 minutos20. Su concentración aumenta paulatinamente durante el día y alcanza un pico alrededor de la medianoche, para decrecer hasta el inicio de un nuevo ciclo, que comenzaría con la aparición de la luz solar. Este patrón depende también de la alimentación. Así, los niveles circulantes de leptina aumentan en las primeras horas después de la ingesta y continúan su ascenso en caso de sobrealimentación. En situaciones de ayuno, hay un descenso en la producción de leptina. Por otro lado, parece que los cambios en el patrón de secreción asociados a la alimentación están más relacionados con la concentración plasmática de insulina que con el peso corporal. Esto se debe a que la insulina estimula la expresión de la leptina en adipocitos aislados y por tanto eleva su nivel circulante.
En la secreción de leptina intervienen también otras hormonas. Los glucocorticoides estimulan su síntesis en adipocitos cultivados, aunque los niveles plasmáticos de cortisol y de leptina conservan una relación inversa a lo largo del tiempo. Esto significa que las concentraciones pico de cortisol coinciden con las concentraciones más bajas de leptina y viceversa20. Aún no se conoce la razón de esta aparente paradoja. Los niveles de leptina también están correlacionados en forma negativa con los de la hormona de crecimiento21, si bien los mecanismos subyacentes no han sido comprendidos a cabalidad. Aunque existen reportes contradictorios, se ha observado que la administración de hormonas tiroideas disminuye la secreción de leptina, mientras que en pacientes hipotiroideos está disminuida22.
Los procesos infecciosos, así como algunas citocinas relacionadas (factor de necrosis tumoral (TNF) e interleucina 1) estimulan la síntesis de leptina23, lo que puede conducir a anorexia y a pérdida del apetito en estados patológicos como el cáncer y las infecciones. Las catecolaminas disminuyen la expresión de la leptina, efecto mediado a través de la activación de receptores b-adrenérgicos. La exposición al frío, el ejercicio exhaustivo y de larga duración (más de 45 minutos) son también condiciones que reducen la leptinemia, hecho que puede interpretarse como una señal hormonal de gasto significativo de energía.
La producción de leptina placentaria es intensamente estimulada tanto por hipoxia como por la insulina y el cortisol. La alimentación, la colecistocinina (CCK) y la gastrina, si bien disminuyen la síntesis de leptina gástrica, aumentan la cantidad de leptina plasmática por medio de mecanismos hasta ahora desconocidos1.
La leptina circula unida con varias proteínas plasmáticas como la albúmina. La proporción de leptina unida con proteínas es más baja en individuos obesos24. El receptor ObRe circula unido con la leptina y funciona como un regulador de la concentración de hormona libre.
La leptina atraviesa la barrera hematoencefálica mediante un mecanismo de transporte asociado con receptor, del mismo modo que ocurre con otras proteínas25. Este mecanismo se explica, si se tiene en cuenta que el endotelio cerebral expresa abundantes receptores para leptina, en especial del subtipo ObRa y que además es capaz de ligar y de internalizar la hormona26. Entre otras cosas, éste parece ser el principal papel de la forma �corta� del receptor. La leptina también puede ser llevada hacia el sistema nervioso central por medio del líquido cefalorraquídeo, donde su concentracion corresponde a una fracción fija de la plasmática. En este tipo de transporte participa también el receptor ObRa, expresado en el plexo coroideo26.
Con una vida media similar en individuos obesos y no obesos, de cerca de 25 minutos27, la leptina es metabolizada en especial por las células epiteliales renales28. Estas internalizan la molécula por medio de un mecanismo mediado por receptores �cortos� (ObR a, c y d), razón por la que su concentración aumenta en pacientes con insuficiencia renal29. El hígado no parece tener un papel importante en el metabolismo de la leptina, aunque se requiere una mayor evidencia.
FUNCIÓN DE LA LEPTINA EN EL CONTROL DEL APETITO Y DEL PESO CORPORAL
Desde su descubrimiento, el papel más estudiado de la leptina es el de reguladora del peso corporal. El órgano blanco involucrado en esta función es el hipotálamo. ObRb presenta un elevado nivel de expresión en neuronas de varios núcleos hipotalámicos, como el arcuado, el paraventricular, los ventromediales y los dorsomediales30, todos implicados en la regulación del comportamiento alimentario y del balance energético. El núcleo arcuado no sólo es el que expresa una mayor cantidad de ObRb sino además, el principal sitio de acción de la leptina en el hipotálamo. Comprende dos poblaciones de neuronas blanco de la leptina: la primera forma parte de la vía orexígenica (inductora del apetito), y está constituida por neuronas que liberan neuropéptido Y (NPY) y por neuronas productoras de AGRP (del inglés AGouti-Related Protein). La segunda forma parte de la vía anorexigénica (inductora de saciedad) y comprende neuronas secretoras de propiomelanocortina (POMC) y de su subproducto aMSH, así como por neuronas que liberan CART (del inglés Cocaine and Amphetamine-Regulated Transcript). En los ratones ob/ob y db/db coexisten niveles aumentados de NPY/AGRP con niveles disminuídos de POMC/CARTY, situación que puede ser modificada en sentido inverso, cuando se administra leptina a los ratones ob/ob.
A partir de estos y otros estudios se infiere que la leptina inhibe la vía orexigénica y estimula la anorexigénica. La leptina también interactúa con otras poblaciones de células reguladoras del apetito, como las neuronas que producen endocanabinoides, hormona concentradora de la melatonina y orexinas31. Las neuronas de las vías orexigénica y anorexigénica inervan varios centros del hipotálamo y del tallo cerebral, de modo que dan origen a las complejas señales integradoras responsables del comportamiento alimentario del individuo.
Los individuos obesos, aunque presentan un alto nivel de leptina desarrollan de forma simultánea una resistencia a la acción de la hormona32, posiblemente a causa del funcionamiento inadecuado de las cascadas de señalización intracelular asociadas con el receptor activado. Este hecho no permite que la hormona ejerza su acción anorexigénica y que además, la leptina per se no tenga utilidad clínica como punta de lanza en el tratamiento contra la obesidad, como se pensó en un comienzo. Este propósito sólo tendría validez en pacientes con obesidad debida a deficiencia de la hormona, los que, como antes se mencionó, son muy escasos.
FUNCIONES METABÓLICAS E INTERACCIÓN CON OTRAS CITOCINAS Y CON LA INSULINA
Las interacciones entre la leptina y la insulina han sido exploradas extensamente, debido por una parte a la coexistencia de estados de resistencia a insulina y a leptina en individuos obesos y por otra, a la clara asociación entre obesidad y diabetes mellitus no insulino dependiente (DMNID). La DMNID se caracteriza por un estado severo de resistencia a la insulina asociado con hiperglicemia, lo que ha permitido postular a la leptina como la responsable de las relaciones entre obesidad y resistencia a la insulina y entre obesidad y DMNID33. La evidencia demuestra que en los adipocitos de rata, la leptina disminuye la unión de la insulina con sus receptores34. Además de su notoria acción lipolítica sobre estas células, la leptina también inhibe los efectos antilipolítico y lipogénico de la insulina35, aunque tal efecto aún no se haya demostrado en adipocitos humanos. La leptina y la insulina se regulan mutuamente. Así, la leptina inhibe la producción de insulina en las células b del páncreas36, mientras que la insulina estimula la producción de leptina en el adipocito. Como antes se mencionó, la leptina y la insulina afectan a las vías de señalización intracelular comunes y a las proteínas IPS, indicio de la importancia de su interacción fisiológica. En el tejido adiposo de roedores23 y del hombre37, se ha demostrado que el factor de necrosis tumoral a (FNTa), citocina producida por las células del sistema inmune y por el tejido adiposo, estimula la expresión y por tanto la producción de la leptina. Por otro lado, la interleucina 1b también disminuye la expresión y la producción de leptina, mientras que las interleucinas 6 y 8 no la afectan38.
FUNCIÓN EN EL CONTROL DEL INICIO DE LA PUBERTAD Y LA REPRODUCCIÓN
El papel de la leptina sobre el inicio de la pubertad es objeto de controversia, debido a que estudios previos han informado resultados contradictorios. La primera indicación acerca de la leptina como agente crítico en el desarrollo puberal surgió de la infertilidad evidente de los ratones ob/ob y db/db y de la correlación entre mutaciones en estos genes con hipogonadismo e infertilidad en otras especies39. En los roedores se produce un pico de secreción de leptina antes de que comience la elevación en el nivel de hormonas sexuales, propio del inicio de la pubertad. Debe anotarse que estos cambios son independientes de la masa corporal y ocurren tanto en machos como en hembras39. En los seres humanos y en otros primates, también se ha informado un aumento en el nivel de leptina un poco antes o, simultáneamente con el inicio de la pubertad40.
El nivel circulante de leptina es mayor en mujeres que en hombres a lo largo de toda la vida, aunque esta diferencia es más notoria después de la pubertad. Esto se debe, en parte, a que en la mujer, los depósitos de grasa tienden a ser mayores que en el hombre, aún después de ajustar las concentraciones de leptina a la cantidad de grasa corporal. En las mujeres, después del inicio de la pubertad, el nivel circulante de leptina aumenta en forma notoria41 posiblemente debido a la acción de los estrógenos. La testosterona en cambio, tiende a disminuir la leptinemia, afirmación consistente con los hallazgos de Ahmed et al.40 quienes observaron un aumento en la leptina con el inicio de la pubertad femenina y una disminución en los varones. La relación conocida entre un peso corporal crítico y el inicio de la pubertad femenina parece estar relacionada directamente con la producción de leptina. En otras palabras, si no existiera una cantidad suficiente de tejido adiposo para producir una cierta cantidad crítica de leptina, la pubertad no ocurriría, fenómeno frecuente en atletas y en mujeres desnutridas con masa grasa muy escasa o ausente41. En los varones por otro lado, la leptina parece estar relacionada con la regulación del proceso eyaculatorio, si se tiene en cuenta la asociación entre la eyaculación prematura y la hipoleptinemia43.
Durante el embarazo, el nivel circulante de leptina materna aumenta progresivamente y alcanza un máximo durante el segundo trimestre. A partir de este momento permanece elevado hasta el momento del parto43. Este aumento en la producción de leptina es también independiente de la aparición de nuevos depósitos de grasa. La producción de leptina placentaria es mayor al inicio del embarazo y disminuye a medida que avanza, lo que sugiere que durante este período, la placenta no es la principal fuente de la hormona, como inicialmente se creyó. El nivel fetal de leptina depende en un alto porcentaje del materno. Por esta razón la leptinemia disminuye abruptamente en el recién nacido.
La leptina puede atravesar la barrera hematoplacentaria por un mecanismo mediado por receptores cortos, similar al utilizado para atravesar la barrera hematoencefálica. Aunque a la leptina en el embarazo se le han asignado funciones como factor de crecimiento y como estimulante de la angiogénesis y de la hematopoyesis fetal44, se requieren más estudios al respecto. Debido a que la producción de leptina está anormalmente aumentada en los estados de preeclampsia45, se ha postulado que la hiperleptinemia es un marcador de insuficiencia placentaria, de utilidad clínica como predictor de las condiciones patológicas relacionadas.
Como antes se mencionó, la leptina es también producida por la glándula mamaria y secretada en la leche. Durante la lactancia, la producción mamaria aumenta notablemente y puede actuar como un factor paracrino estimulante de la proliferación del tejido mamario y de la producción láctea46.
El nivel circulante de leptina en el recién nacido está correlacionado en forma positiva con parámetros tales como peso, talla, perímetro cefálico, superficie corporal y edad gestacional47. Durante la niñez, la leptinemia continúa en un nivel bajo hasta la pubertad, de modo dependiente de la cantidad de grasa corporal.
FUNCIÓN COMO FACTOR DE CRECIMIENTO Y RELACIÓN CON EL CÁNCER
La leptina actúa como mitógeno de todas las células estudiadas, p.e, . las células endoteliales48, las células musculares lisas49, las células epiteliales normales y las neoplásicas de colon50 y mama51. La hormona es especialmente importante durante el proceso de angiogénesis, tanto en tejidos normales como neoplásicos, hecho demostrado in vivo e in vitro. Bouloumie48 informó que la leptina aumenta la tasa de proliferación vascular endotelial, con una eficiencia similar a la del factor de crecimiento vascular (VEGF). Otros estudios han mostrado el poderoso efecto estimulante de la leptina sobre el crecimiento, la proliferación y la migración de las células endoteliales, así como la acción promotora sobre la expresión de varios genes involucrados en la angiogénesis. Es así como estudios preliminares han correlacionado un nivel elevado de leptina con la aparición de diversos tipos de cáncer52. Posiblemente este es el fundamento molecular de las asociaciones epidemiológicas entre la obesidad y el cáncer, establecidas de forma fehaciente en cáncer de seno y de colon. Con relación al cáncer de próstata se ha demostrado que niveles aumentados de leptina inciden no sólo sobre el desarrollo de malignidad53 sino sobre un peor pronóstico y enfermedad extraprostática, cuando la lesión maligna ya está establecida54.
La hiperleptinemia concurre con un mayor riesgo de cáncer de seno55. En este sentido, parece existir una sinergia con los estrógenos, porque la leptina induce actividad en la aromatasa, enzima encargada de transformar otros esteroides en estrógenos dentro del tejido mamario. Adicionalmente, se han encontrado receptores para la leptina en las células cancerosas de seno. Como éstas a su vez poseen receptores estrogénicos la hormona puede estimular su proliferación51. De manera semejante, se ha observado que la leptina estimula la actividad del sistema JAK-STAT356 en las células cancerosas y que existe relación entre niveles elevados de leptina y cáncer, tanto endometrial como del cuello uterino57.
Por otro lado, ha sido demostrado que la leptina promueve el crecimiento de las células epiteliales colónicas normales y malignas50, lo que concuerda con la presencia de receptores hormonales expresados a todo lo largo del tracto gastrointestinal59. En este sentido, se dispone de evidencia sobre la relación existente entre hiperleptinemia y carcinomas de colon60 y de esófago61. En el estómago aumenta la expresión de la leptina cuando hay infección con la bacteria Helicobacter pylori, reconocida por su potencial cancerígeno62. También se ha sugerido que el consumo de fibra brinda protección contra el cáncer de colon, debido a la disminución de la producción de leptina. De modo inverso, cuando aumenta la producción de la hormona, aparece una predisposición al desarrollo de cáncer de colon, causada por el consumo de grasas saturadas63.
En varias líneas celulares leucémicas linfoides y mieloides, ha sido identificado el ARNm de algunos subtipos del receptor de leptina, lo que explica que la hormona pueda estimular la proliferación de dichas células tanto in vivo como in vitro64. También se ha propuesto una posible relación entre la hiperleptinemia y el cáncer de vejiga65.
LEPTINA Y FUNCIÓN INMUNE
La leptina es un modulador de la respuesta inmune, de manera semejante a lo que ocurre con otras citocinas. Cuando actúa sobre los receptores de células mononucleares y linfoides estimula la producción de citocinas proinflamatorias66. Si lo hace sobre los receptores ObRb expresados en las líneas celulares NK92 (natural killer) derivadas de linfocitos, la leptina estimula no sólo su proliferación sino también su actividad citotóxica contra células tumorales67. En adición, la leptina puede actuar como factor estimulante de los linfocitos T ayudadores68. Debido a sus propiedades inmunoestimulantes, se ha planteado que la hipoleptinemia es la causa de la disfunción inmune asociada con desnutrición y con estados inductores de caquexia68.
FUNCIÓN EN EL DESARROLLO Y ACTIVIDAD DEL SNC
En estudios recientes se ha demostrado que LR se expresa en otras áreas del cerebro no relacionadas directamente con el control alimentario, tales como cerebelo, corteza piriforme, corteza cerebral, tálamo, hipocampo, amígdala, tracto olfatorio y sustancia nigra69, donde la función de la leptina es objeto de estudio.
En el hipocampo, área crítica para el aprendizaje y la memoria, la leptina favorece la potenciación a corto y a largo plazo, mediada por receptores NMDA de glutamato, mediante mecanismos relacionados con la activación de PI3 kinasa70. Así, los ratones db/db muestran no sólo notables alteraciones de la potenciación sino depresión a largo plazo, lo que incide en su proceso de aprendizaje71.
ASOCIACIONES DE LEPTINA Y ENFERMEDADES PSIQUIÁTRICAS
Desde hace varios años, los investigadores han intentado relacionar las enfermedades psiquiátricas con la leptina, motivados quizás por las asociaciones establecidas previamente entre algunas de ellas y la obesidad. Algunos estudios han hallado relación entre la hiperleptinemia y entidades tales como trastorno de personalidad limítrofe72, esquizofrenia e intento de suicidio73. Otros han vinculado la hipoleptinemia con la depresión74 y con el síndrome de estrés postraumático persistente75. Aunque los significados fisiopatológico y clínico de tales relaciones aún no está establecido con claridad, la evidencia plantea interesantes premisas a evaluar en investigaciones futuras.
LEPTINA, HUESO Y CARTÍLAGO
Los osteoblastos y los condrocitos expresan ObRb76, indicio de la actividad de la hormona dentro de estas células. En roedores77 y en seres humanos78 la leptina aumenta la masa mineral ósea, mientras que in vitro79 promueve la proliferación y el crecimiento de osteoblastos y condrocitos. Los efectos osteogénicos mencionados son mediados por acciones periféricas directas sobre los osteoblastos. Por otro lado, los ratones ob/ob tienen una mayor densidad ósea, normalizada por administración de leptina80, lo que sugiere que la hormona tiene efectos antiosteogénicos, en apariencia mediados por mecanismos centrales hipotalámicos. Aunque la naturaleza exacta de tales mecanismos aún no ha sido dilucidada, se sabe que estimulan al sistema simpático, el que a su vez actúa sobre los receptores b2 adrenérgicos de la membrana del osteoblasto, con la finalidad de reducir la formación ósea80. En adición, Dumond et al.81 informaron aumentos en la producción de leptina y en la expresión de sus receptores, en condrocitos de cartílago osteoartrítico, lo que demostró la participación de la hormona en la fisiopatología de la enfermedad.
LEPTINA Y SISTEMA CARDIOVASCULAR
Algunos investigadores han postulado que la hiperleptinemia es la explicación de la relación entre la obesidad y las enfermedades cardiovasculares82. La leptina produce un aumento significativo de la actividad simpática, gracias a su acción central sobre el hipotálamo, posiblemente debido a que estimula la liberación de melanocortina y de la hormona liberadora de corticotropina (CRH). En roedores, la inyección intraventricular de leptina eleva la presión arterial media y la frecuencia cardíaca y aumenta además la actividad simpática renal en forma significativa83. Se han observado efectos semejantes cuando la hormona es administrada en forma intravenosa, aunque en menor magnitud y más tardíamente84. Estas acciones pueden ser inhibidas con la administración previa de bloqueadores simpáticos, lo que indica la mediación del sistema simpático. Sin embargo, los efectos periféricos de la leptina son totalmente opuestos, pues la hormona estimula la liberación de óxido nítrico (NO), potente vasodilatador local, por parte de las células endoteliales85.
Así pues, la leptina es un posible regulador del tono vascular y por ende de la presión arterial, mediante sus efectos centrales sistémicos y periféricos locales. Aunque dichos efectos en apariencia se contrarrestan, pueden llegar a complementarse dentro del proceso de regulación global. Este hecho tendría relevancia en la patogénesis de la hipertensión arterial, que ha sido usualmente asociada con la hiperleptinemia82,86.
CONCLUSIONES
Si bien la investigación sobre la leptina ha avanzado en los últimos años, lo que ha permitido establecer su relación con múltiples procesos fisiológicos y patológicos, el conocimiento acerca de sus funciones e interacciones con otras hormonas aún es limitado. Con absoluta certeza se sabe de su papel en el control del peso corporal y de una gran diversidad de procesos, que comprenden funciones neuroendocrinas complejas tales como metabolismo, crecimiento, reproducción, función inmune y tono vascular, entre otras. La investigación presente y futura tiene la misión de responder a la gran cantidad de interrogantes sobre esta hormona y de establecer cuál es su verdadero papel en la fisiopatología del amplio rango de enfermedades con las que ha sido vinculada, como cáncer, la DMNID y enfermedades cardiovasculares.
REFERENCIAS
1. Ahima RS, Flier JS. Leptin. Ann Rev Physiol 2000; 62: 413-437.
2. Harris RB, Kasser TR, Martin RJ. Dynamics of recovery of body composition after overfeeding, food restriction or starvation of mature female rats. J Nutr 1986; 116: 2536-2546.
3. Hetherington AW, Ranson SW. Hypothalamic lessions and adiposity in the rat. Anat Rec 1940; 78: 149-172.
4. Hervey GR. The effects of lesions in the hypothalamus in parabiotic rats. J Physiol 1958; 145: 336-352.
5. Ingalls AM, Dickie MM, Snell GD. Obesity, a new mutation in the house mouse. J Hered 1950; 41: 317-318.
6. Coleman DL. Effects of parabiossis of obese with diabetes and normal mice. Diabetologia 1973; 9: 294-298.
7. Zhang Y, Proenca R, Maffei M, Barone L, Leopold L, Friedman JM. Positional cloning of the mouse obese gene and its human homologue. Nature 1994; 372: 425-432.
8. Montague CT, Farooqui S, Whitehead JP, et al. Congenital leptin deficiency is associated with severe early onset obesity in humans. Nature 1997; 387: 903-908.
9. Fei H, Okano HJ, Li C, et al. Anatomic localization of alternatively spliced leptin receptors (Ob-R) in mouse brain and other tissues. Proc Natl Acad Sci USA 1997; 94: 7001-7005.
10. Clement K, Vaisse C, Lahlous N, et al. A mutation in the human leptin receptor gene causes obesity and pituitary dysfunction. Nature 1998; 392: 398-401.
11. Lee GH, Proenca R, Montez JM, et al. Abnormal splicing of the leptin receptor in diabetic mice. Nature 1996; 379: 632-635.
12. Bjorbaek C, Uotani S, Da Silva B, Flier JS. Divergent signaling capacities of the long and short isoforms of the leptin receptor. J Biol Chem 1997; 272: 32686-32695.
13. Ghilardi N, Skoda RC. The leptin receptor activates Janus tyrosine kinase 2 and signals for proliferation in a factor-dependent cell line. Molecular Endocrinol 1997; 11: 393-399.
14. Myers MG Jr, White MF. The molecular basis of insulin action. In: DeGroot, LJ, Jameson JL (eds.). Endocrinology. 4th ed. Philadelphia: Saunders. p. 712-727.
15. Spanswick D, Smith MA, Groppi V, Logan SD, Ashford ML. Leptin inhibits hypothalamic neurons by activation of ATP-sensitive potassium channels. Nature 1997; 390: 521-525.
16. Harvey J, McKenna F, Herson PS, Spanswick D, Ashford ML. Leptin activates ATP-sensitive potassium channels in the rat insulin-secreting line, CRI-G1. J Physiol 1997; 504: 527-535.
17. Shanley LJ, Irving AJ, Rae MG, Ashford ML, Harvey J. Leptin inhibits rat hippocampal neurons via activation of large conductance calcium-activated K+ channels. Nat Neurosci 2002; 5: 299-300.
18. Prins JB. Adipose tissue as an endocrine organ. Best Prac Res Clin Endocrinol Metab 2002; 16: 639-651.
19. Masuzaki H, Ogawa Y, Sagawa N, et al. Nonadipose production of leptin: leptin as a novel placenta-derived hormone in humans. Nat Med 1997; 3: 1029-1033.
20. Licinio J, Mantzoros C, Negrao AB, et al. Human leptin levels are pulsatile and inversely related to pituitary-adrenal function. Nat Med 1997; 3: 575-579.
21. Ghizzoni L, Mastorakos G. Interactions of leptin, GH and cortisol in normal children. Ann NY Acad Sci 2003; 997: 56-63.
22. Zimmermann-Belsing T, Brabant G, Holst JJ, Feldt-Rasmussen U. Circulating leptin and thyroid dysfunction. Eur J Endocrinol 2003; 149: 257-271.
23. Sarraf P, Frederich RA, Turner EM, et al. Multiple cytokines and acute inflammation raise mouse leptin levels: potential role in inflammatory anorexia. J Exp Med 1997; 185: 171-175.
24. Sihna MK, Ohannesian JP, Heiman ML, et al. Nocturnal rise of leptin in lean, obese and non-insulin dependent diabetes mellitus subjects. J Clin Inv 1996; 97: 1344-1347.
25. Banks WA, Kastin AJ, Huang W, Jaspan JP, Maness LM. Leptin enters the brain by a saturable system independent of insulin. Peptides 1996; 17: 305-311.
26. Bjorbaek C, Elmquist JK, Michl P, et al. Expression of leptin receptor isoforms in brain microvessels. Endocrinol 1998; 139: 3485-3491.
27. Klein S, Coppack V, Mohamed-Ali V, Landt M. Adipose tissue leptin production and plasma leptin kinetics in humans. Diabetes 1996; 45: 984-987.
28. Meyer C, Robson D, Rackovsky N, et al. Role of the kidney in human leptin metabolism. Am J Physiol 1997; 273: E903-E907.
29. Merabet E, Dagogo-Jack S, Coyne DW, et al. Increased plasma leptin concentration in end-stage renal disease. J Clin Endocrinol Metab 1997; 82: 847-850.
30. Elmquist JK, Bjorbaek C, Ahima RS, Flier JS, Saper CB. Distributions of leptin receptor mRNA isoforms in the rat brain. J Comp Neurol 1998; 395: 535-547.
31. Harvey J, Ashford ML. Leptin in the CNS: much more than a satiety signal. Neuropharmacol 2003; 44: 845-854.
32. Friedman JM, Halaas JL. Leptin and the regulation of body weight in mammals. Nature 1998; 395: 763-770.
33. Ceddia RB, Heikki AK, Zierath JR, Sweeney G. Analysis of paradoxical observations on the association between leptin and insulin resistance. FASEB J 2002; 16: 1163-1176.
34. Walder K, Filippis A, Clark S, Zimmer P, Collier GR. Leptin inhibits insulin binding in isolated rat adipocytes. J Endocrinol 1997; 155: R5-R7.
35. Rodríguez VM, Macarulla MT, Echevarría E, Portillo MP. Lipolysis induced by leptin in rat adipose tissue from different anatomical locations. Eur J Nutr 2003; 42: 149-153.
36. Seufert J. Leptin effects on pancreatic beta-cell gene expression and function. Diabetes 2004; 53 Supl. 1: 152-158.
37. Zumbach MS, Boehme MW, Wahl P, Stremmel W, Ziegler R, Nawroth PP. Tumor necrosis factor increases serum leptin levels in humans. J Clin Endocrinol Metabolism 1997; 82: 4080-4082.
38. Bruun JM, Pedersen SB, Kristensen K, Richelsen B. Effects of pro-inflammatory cytokines and chemokines on leptin production in human adipose tissue in vitro. Mol Cell Endocrinol 2002; 190: 91-99.
39. Goumenou AG, Matalliotakis IM, Koumantakis GE, Panidis DK. The role of leptin in fertility. Eur J Obst Gynecol Reprod Biol 2003; 106: 118-124.
40. Ahmed ML, Ong KK, Morrell DJ, et al. Longitudinal study of leptin concentrations during puberty: sex differences and relationship to changes in body composition. J Clin Endocrinol Metab 1999; 84: 899-905.
41. Apter D. The role of leptin in female adolescence. Ann NY Acad Sci 2003; 997: 64-76.
42. Atmaca M, Kuloglu M, Tezcan E, Semercioz A, Ustundag B, Ayar A. Serum leptin levels in patients with premature ejaculation. Arch Androl 2002; 48: 345-350.
43. Lage M, García-Mayor RV, Tome MA, et al. Serum leptin in women throughout pregnancy and the postpartum period and in women suffering spontaneous abortion. Clin Endocrinol 1999; 50: 211-216.
44. Henson MC, Castracane D. Leptin in pregnancy. Biol Reprod 2000; 63: 1219-1228.
45. Anderson CM, Ren J. Leptin, leptin resistance and endothelial dysfunction in pre-eclampsia. Cell Mol Biol 2002; 48: 323-329.
46. Bonnet M, Delavaud C, Laud K, et al. Mammary leptin synthesis, milk leptin and their putative physiological roles. Reprod Nutr Dev 2002; 42: 399-413.
47. Su PH, Wang SL, Chen JY, Lai CP, Jian SH. Serum leptin levels in preterm, healthy and sick-term newborns. Acta Pediatr Taiwan 2002; 43: 249-254.
48. Boulomie A, Drexler HC, Lafontan M, Busse R. Leptin, the product of Ob gene promotes angiogenesis. Circ Res 1998; 83: 1059-1066.
49. Oda A, Taniguchi T, Yokoyama M. Leptin stimulates rat aortic smooth muscle cell proliferation and migration. Kobe J Med Sci 2001; 47:141-150.
50. Hardwick JC, Van Den Brink GR, Offerhaus GJ, Van Deventer SJ, Peppelenbosch MP. Leptin is a growth factor for colonic epithelial cells. Gastroenterol 2001; 121: 79-90.
51. Dieudonne MN, Machinal-Quelin F, Serazin-Leroy V, Leneveu MC, Pecquery R, Giudicelli Y. Leptin mediates a proliferative response in human MCF7 breast cancer cells. Biochem Biophys Res Commun 2002; 293: 622-628.
52. Somasundar P, MsFadden DW, Hileman SM, Vona-Davis L. Leptin is a growth factor in cancer. J Surg Res 2004; 116: 337-349.
53. Stattin P, Soderberg S, Hallmans G, et al. Leptin is associated with increased prostate cancer risk: a nested case-referent study. J Clin Endocrinol Metab 2001; 86: 1341-1345.
54. Chang S, Hursting SD, Contois JH, et al. Leptin and prostate cancer. Prostate 2001; 46: 62-67.
55. Tessitore L, Vizio B, Pesola D, et al. Adipocyte expression and circulating levels of leptin increase in both gynaecological and breast cancer patients. Int J Oncol 2004; 24: 1529-1535.
56. Catalano S, Mauro L, Marsico S et al. Leptin induces, via ERK1/ERK2 signal, functional activation of estrogen receptor alpha in MCF-7 cells. J Biol Chem 2004; 279: 19908-19915.
57. Petridou E, Belechri M, Dessypris N, et al. Leptin and body mass index in relation to endometrial cancer risk. Ann Nutr Metab 2002; 46: 147-151.
58. Yuan SS, Tsai KB, Chung YF, et al. Aberrant expression and possible involvement of the leptin receptor in endometrial cancer. Gynecol Oncol 2004; 92: 769-775.
59. Morton NM, Emilsson V, Liu YL, Cawthorne MA. Leptin action in intestinal cells. J Biol Chem 1998; 273: 26194-26201.
60. Stattin P, Palmqvist R, Soderberg S, et al. Plasma leptin and colorectal cancer risk: a prospective study in Northern Sweden. Oncol Rep 2003; 10: 2015-2021.
61. Somasundar P, Riggs D, Jackson B, Vona-Davis L, McFadden DW. Leptin stimulates esophageal adenocarcinoma growth by nonapoptotic mechanisms. Am J Surg 2003;186: 575-578.
62. Azuma T, Suto H, ItoY, et al. Gastric leptin and Helicobacter pylori infection. Gut 2001; 49: 324-329.
63. Lin X, Chávez MR, Bruch RC, et al. The effects of a high fat diet on leptin mRNA, serum leptin and the response to leptin are not altered in a rat strin susceptible to high fat diet-induced obesity. J Nutr 1998; 128: 1606-1613.
64. Cioffi JA, Shafer AW, Zupandic TJ, et al. Novel B219/OB receptor isoforms: possible role of leptin in hematopoiesis and reproduction. Nat Med 1996; 2: 585-589.
65. Yuan SS, Chung YF, Chen HW, et al. Aberrant expression and possible involvement of the leptin receptor in bladder cancer. Urology 2004; 63: 408-413.
66. Sánchez-Margalet V, Martín-Romero C, Santos-Alvarez J, Goberna R, Najib S, González-Yanes C. Role of leptin as an immunomodulator of blood mononuclear cells: mechanisms of action. Clin Exp Immunol 2003; 133: 11-19.
67. Zhao Y, Sun R, You L, Gao C, Tian Z. Expression of leptin receptors and response to leptin stimulation of human natural killer cell lines. Biochem Biophys Res Commun 2003; 300: 247-252.
68. Matarese G, Sanna V, Fontana S, Zappacosta S. Leptin as a novel therapeutic target for immune intervention. Curr Drug Targets Inflamm Allergy 2002; 1: 13-22.
69. Hakansson MM, Brown H, Guilardi N, Skoda RC, Meister BJ. Leptin receptor immunoreactivity in chemical-defined target neurons of the hypothalamus. J Neurosci 1998; 18: 559-572.
70. Shanley LJ, Irving AJ, Harvey J. Leptin enhances NMDA receptor function and modulates hippocampal synaptic plasticity. J Neurosci 2001; 186: 1-6.
71. Li XL, Aou S, Oomura Y, Hori N, Fukunaga K, Hori T. Impairment of long-term potentiation and spacial memory in leptin receptor-deficient rodents. Neurosci 2002; 113: 607-615.
72. Atmaca M, Kuloglu M, Tezcan E, Gecici O, Ustundag B. Serum cholesterol and leptin levels in patients with borderline personality disorder. Neuropsychobiol 2002; 45: 167-171.
73. Atmaca M, Kuloglu M, Tezcan E, Ustundag B. Serum leptin and cholesterol levels in schizophenic patients with and without suicide attempts. Acta Psychiatr Scand 2003; 108: 208-214.
74. Miller GE, Freedland KE, Carney RM, Stetler CA, Banks WA. Pathways linking depression, adiposity and inflammatory markers in healthy young adults. Brain Behav Immun 2003; 17: 276-285.
75. Liao SC, Lee MB, Lee YJ, Huang TS. Hyperleptinemia in subjects with persistent partial posttraumatic stress disroder after a major earthquake. Psychosom Med 2004; 66: 23-28.
76. Figenschau Y, Knutsen G, Shahazeydi S, Johansen O, Sveinbjornsson B. Human articular chondrocytes express functional leptin receptors. Biochem Biophys Res Commun 2001; 287: 190-197.
77. Steppan CM, Crawford DT, Chidsey-Frink KL, Ke H, Swick AG. Leptin is a potent stimulator of bone growth in ob/ob mice. Regulatory Peptides 2000; 92: 73-78.
78. Thomas T, Burguera B, Melton LJ, et al. Role of serum leptin, insulin and estrogen levels as potential mediators of the relationships between fat mass and bone mineral density in men versus women. Bone 2001; 29: 114-120.
79. Maor G, Rochwerger M, Segev Y, Phillip M. Leptin acts as a growth factor on the chondrocytes of skeletal growth centers. J Bone Miner Res 2002; 17: 1034-1043.
80. Takeda S, Elefteriou F, Levasseur R, et al. Leptin regulates bone formation through a hypotalamic relay: a central control of bone mass. Cell 2002; 11: 305-317.
81. Dumond H, Presle N, Terlain B, et al. Evidence for a key role of leptin in osteoarthritis. Arthritis Rheum 2003; 48: 3118-3129.
82. Sowers JR. Obesity as a cardiovascular risk factor. Am J Med 2003; 115: 37S-41S.
83. Correia ML, Morgan DA, Sivitz WI, Mark AL, Haynes WG. Leptin acts in the central nervous system to produce dose-dependent changes in arterial pressure. Hypertension 2001; 37: 936-942.
84. Shek EW, Brands MW, Hall JE. Chronic leptin infusion increases arterial pressure. Hypertension 1998; 31: 409-414.
85. Frünbeck G. Pivotal role of nitric oxide in the control of blood pressure after leptin administration. Diabetes 1999; 48: 903-908.
86. Coatmellec-Taglioni G, Ribiere C. Factors that influence the risk of hypertension in obese individuals. Curr Opin Nephrol Hypertens 2003; 12: 305-308.