
Eikenella corrodens: Patogénesis y aspectos clínicos*
Rubén Darío Jaramillo, M.Sc., Ph.D.1, Paola Suárez, M.Sc.2, Beatriz Barraza, M.Sc.3, Paulina Lara, M.Sc., Ph.D.4, Luis Teherán, Microbiol.5, José Edgardo Escamilla, M.Sc., Ph.D.6
* Trabajo financiado por Colciencias y la Universidad Libre, Seccional Barranquilla, Colombia, Código 1209-05-10141 CT 267-2000.
1. Director Grupo de Investigación de Biomembranas (GIBIOM), Programa de Medicina, Facultad Ciencias de la Salud, Centro de Investigación (CIUL), Universidad Libre, Seccional Barranquilla, Colombia. e-mail: rubenjaramillo@hotmail.com
2. Co-Investigadora invitada, Grupo GIBIOM, Facultad de Medicina, Universidad de Cartagena, Colombia. e-mail: paolains@hotmail.com
3. Investigadora y Docente, Grupo GIBIOM, Facultad Ciencias de la Salud, CIUL, Universidad Libre, Seccional Barranquilla, Colombia. e-mail: beatriz_soil@yahoo.es
4. Investigadora, Docente, Grupo GIBIOM, Facultad Ciencias de la Salud, CIUL, Universidad Libre, Seccional Barranquilla, Colombia. e-mail: plaracal@gmail.com
5. Coinvestigador, Grupo GIBIOM, Facultad Ciencias de la Salud, CIUL, Universidad Libre, Seccional Barranquilla, Colombia. e-mail: ltheran@unilibrebaq.edu.co
6. Investigador Titular, Departamento de Bioquímica, Instituto de Fisiología Celular, Universidad Nacional Autónoma de México (UNAM), México. e-mail: eescami@ifc.unam.mx
Recibido para publicación agosto 9, 2004 Aceptado para publicación junio 15, 2006
RESUMEN
El ambiente microbiológico oral es único y tiene una dinámica compleja. Se calcula que cerca de 500 especies de bacterias habitan la cavidad oral humana, y alrededor de 22 géneros son los predominantes. Las bacterias que se aíslan con más frecuencia de los sitios infectados de la cavidad oral, y que son también patógenos potenciales, forma un grupo pequeño de microorganismos gramnegativos, entre los que se incluyen los siguientes: Actinobacillus actinomycetemcomitans, Bacteroides forsythus, Campylobacter spp., Capnocytophoga spp., Eikenella corrodens, Fusobacterium nucleatum, Porphyromonas gingivalis, Prevotella intermedia y el grupo-Streptococcus milleri. E. corrodens se reconoce como un microbio patógeno oportunista en la cavidad oral; también, puede causar infecciones extra orales, como un agente infeccioso solo o en combinaciones con diversas bacterias. La presente revisión, con un énfasis en E. corrodens, mostrará varios aspectos de sus características microbiológicas y bioquímicas, también se sistematizan y discuten el conocimiento actual sobre los mecanismos de patogénesis, como los lipopolisacáridos, proteínas externas de membrana, complejo de adesina, pili y otros; infecciones oral y extra oral; factores predisponentes; endocarditis; osteomielitis, infecciones intra-abdominales; diagnóstico y agentes antimicrobianos.
Palabras clave: Eikenella corrodens; Patogénesis; Infecciones extra-orales; Diagnóstico; Antimicrobiano.
Eikenella corrodens: Pathogenesis and clinic aspects
SUMMARY
The oral microbiological environment is unique and has a complex dynamic. It has been estimated that about 500 species of bacteria inhabit the human oral cavity and about 22 genera of them are the most dominant. The most frequently isolated from infected pockets of the oral cavity bacteria which are also potential pathogens form a small group of gramnegative microorganisms among which are included the following: Actinobacillus actinomycetemcomitans, Bacteroides forsythus, Campylobacter spp., Capnocytophoga spp., Eikenella corrodens, Fusobacterium nucleatum, Porphyromonas gingivalis, Prevotella intermedia, and Streptococcus milleri-group. E. corrodens is distinguished for being an opportunistic oral pathogen. It can also produce extra-oral infections acting either as an infectious agent itself or together with other bacteria. Referring to E. corrodens the present review will not only demonstrate different aspects about microbiological and biochemical characteristics, but also the actual knowledge on pathogenesis mechanism, like polysaccharides, outer membrane protein, lectin-like adhesin complex, pili, and other mechanisms will be systematized and discussed. Oral and extra-oral infections and other risking factors like endocarditis; osteomyelitis, intra-abdominal infections, their diagnosis and antimicrobial agents will be also considered in this paper.
Key words: Eikenella corrodens; Pathogenesis; Extra-oral infections; Diagnostics; Antimicrobial.
Eikenella corrodens, una bacteria gramnegativa, es un habitante frecuente de la cavidad oral y tracto respiratorio superior de los seres humanos, con un comportamiento patógeno oportunista, que ha despertado gran interés en las ciencias biomédicas, básicas y clínicas, por sus mecanismos moleculares de patogénesis y la dificultad para lograr relacionarlo como agente causal, en la práctica clínica, con las enfermedades que puede producir debido a su crecimiento fastidioso; por lo que se hace necesario revisar la literatura sobre esta proteobacteria para establecer las enfermedades de base o los factores predisponentes que conduzcan a la presentación de estas entidades, así como los métodos disponibles para su diagnóstico y los antibióticos más apropiados para su tratamiento.
HISTORIA
Desde 1958, se describió un bacilo gramnegativo aislado de la saliva o de abscesos actinomicóticos que crecía bajo condiciones anaeróbicas o se adaptaba al crecimiento aeróbico sólo después de un número de subcultivos, por lo cual se le clasificó en el género Bacteroides con el nombre de B. corrodens1,2. Luego, los resultados1 demostraron que el crecimiento aeróbico de este microorganismo podía depender de la constitución del medio, sobre todo del contenido de hemina; además, el porcentaje de guanina más citosina (G+C) de las cepas facultativas en 57% a 58%, era bastante diferente del valor aceptado para las especies de Bacteroides; por último, se identificó a la cepa facultativa1 como una nueva especie Eikenella corrodens (ATCC 23834; NCTC 10596), y a la cepa anaeróbica estricta se la denominó B. ureolyticus2.
CARACTERÍSTICAS MICROBIOLÓGICAS Y BIOQUÍMICAS
E. corrodens es un cocobacilo gramnegativo anaeróbico facultativo de crecimiento fastidioso con 0.3 a 0.4 por 1.5 a 4mm (Cuadro 1), pertenece al genero Eikenella de la familia Neisseriaceae de la subdivisiónb proteobacterias; también representan a esta familia los géneros Neisseria, Alysiella, Aquaspirillum, Kingella, Microvirgula, Simonsiella, Vogesella y el grupo Chromobacterium; además, es un habitante normal de la cavidad oral, del aparato respiratorio superior y de la superficie de la mucosa de los sistemas intestinal y genital, con un carácter de agente patógeno oportunista que se ha asociado con infecciones orales y no orales usualmente como parte de un consorcio microbiano. Por lo regular los microorganismos aparecen unidos, son rectos, con final redondeado, no ramificados, no flagelados y no esporulados1 y frente a la tinción de Gram dan reacción negativa. Las colonias sobre agar sangre a las 24 y 48 horas tienen 0.5 mm y 0.5 a 1.0 mm de diámetro, respectivamente; asimismo, las cepas con morfología corroen, piquetean el agar y la colonia aparece rugosa, con bordes circulares o irregulares, con un color gris, translúcida y no hemolítica, algunas veces presenta un halo verdoso pequeño después de 48 horas o más de incubación, y el olor de los cultivos recuerda a los géneros Pasteurella y Haemophilus. El crecimiento en medio líquido es usualmente pobre, pero crece mejor en medios semisólidos y se ve favorecido en presencia de CO2 del 5% al 10%, de manera especial sobre aislamiento primario y con frecuencia en la subsiguiente transferencia para muchos subcultivos. El mejor crecimiento se obtiene en un medio líquido con concentraciones de 5 a 25 mg de hemina/litro de medio y agar 0.2%. Las variantes que no requieren hemina se pueden seleccionar in vitro1.
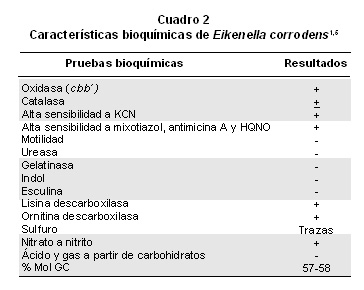
La caracterización bioquímica (Cuadro 2) de E. corrodens señala una bacteria con actividad positiva para oxidasa, lisina descarboxilasa, ornitina descarboxilasa, prolina iminopeptidasa, carboxipeptidasa para glutamato, azocaseinasa, fosfatasa alcalina y ácida, fosfoamidasa, esterasa y esterasa de lípidos3; también, tiene actividad de catalasa variable, trazas de sulfuro de hidrógeno (H2S); pero es ureasa, indol, gelatinasa y esculina negativa. Se caracteriza por ser una bacteria asacarolítica que no produce ácido ni gas a partir de glucosa, xilosa, manitol, lactosa, sacarosa y maltosa, y reduce el nitrato a nitrito1. La producción de energía de E. corrodens parece estar ligada, en forma parcial o total a la desaminación oxidativa de prolina, glutamato, serina y glutamina donde el nitrato actúa como último aceptor de electrones4. Su sistema respiratorio aeróbico está constituido por NADH, succinato y formato deshidrogenasas, ubiquinona, complejo bc1 inhibido por antimicina A, HQNO y mixotiazol, y una citocromo oxidasa cbb´ sensible a cianuro5. El porcentaje por mol de GC de ADN es de 57% a 58%1. Lo anterior sugiere, que se requiere profundizar aún mucho más en el metabolismo catabólico, anabólico y bioenergético.
MECANISMOS DE PATOGÉNESIS
Cuando se relaciona un microorganismo con determinadas entidades clínicas se deben establecer los mecanismos por medio de los cuales es capaz de producir la enfermedad; es decir, los mecanismos de patogenicidad que se relacionan en forma directa con sus componentes estructurales, interacciones entre la misma especie y la presencia de un consorcio microbiano, lo que será útil no sólo para establecer si este microorganismo es realmente el agente causal de la enfermedad, sino también para aplicar una terapéutica eficaz. Entre los componentes de la superficie de E. corrodens que se pueden considerar como factores de virulencia (Cuadro 3), se encuentran los lipopolisácaridos, exopolisacáridos, proteínas de membrana externa, limo, adesinas y pilis2, 6.
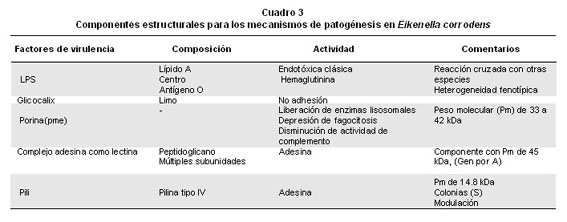
Lipopolisacáridos. Los lipopolisacáridos (LPS) de E. corrodens7 están formados de carbohidratos 34.5% p/p, lípido A 25% p/p y tienen una pequeña cantidad del ácido 2-ceto-3-deoxioctonoico (KDO), heptosa y una fuerte actividad endotóxica clásica8; también, en el estudio de la estructura de los LPS en presencia de anticuerpos monoclonales 8 se encontró un epítope central común entre E. corrodens y Fusobacterium nucleatum, una reacción cruzada entre el epítope O de E. corrodens con F. necrophorum y Capnocytophaga ochracea; mientras, que el lípido A de E. corrodens comparte un epítope común entre los LPS de varios bacilos gramnegativos. Se diferenciaron 16 diferentes fenotipos de LPS entre 27 cepas de E. corrodens9 que fueron examinadas, lo que indica la apreciable heterogeneidad estructural en los LPS.
Los resultados señalan que E. corrodens presenta un factor de hemoaglutinación estable al calor para eritrocitos de ovejas, ratones y humanos, que no es inhibido por galactosa o lactosa10; es decir, no sería semejante a lectina, lo cual se evidencia por los patrones de SDS-PAGE que revelan que esta actividad puede ser causada por lipopolisácaridos de tipo rugoso y parece jugar un papel importante en la unión al área subgingival10. Igualmente, los LPS de E. corrodens son potencialmente mitogénicos para células esplénicas de ratón, induce la resorción del hueso en cultivo de órganos y tiene actividad en el ensayo del lisado de amebocitos del Limulus7.Los resultados anteriores sobre las diferencias en la composición y actividad biológica de los LPS de E. corrodens, indican que se deben considerar como factores potenciales de virulencia asociados con la placa subgingival y la enfermedad periodontal.
Limo. Estudios de microscopía electrónica han demostrado en E. corrodens11 una capa de limo fibroso asociado con la superficie externa de la membrana externa, constituida por exopolisacáridos2. El material de limo amorfo11 aparece recubriendo células en variantes grandes y pequeñas de la cepa silvestre (ATCC 23834). El efecto del extracto de limo en los ensayos de Shwartzman, letalidad en embriones de pollo, lisado de amebocitos del Limulus y mitogenicidad en las células del bazo está bastante disminuido con poca actividad endotóxica, pero parece tener una importante respuesta inmunosupresiva12 pues suprime la respuesta inmune del ratón contra eritrocitos de cordero; finalmente, el potencial antifagocítico de la capa de limo no se ha determinado.
Proteínas de la membrana externa. El análisis por electroforesis en gel de poliacrilamida-dodecil sulfato de sodio (SDS-PAGE) de membrana externa de E. corrodens muestra de una a tres bandas de proteínas principales con peso molecular aparente entre 33 y 43 kDa2,6,9, y una a dos bandas más pequeña entre 24 y 28 kDa9; además, al parecer una proteína de peso molecular de 42 kDa muestra actividad antigénica9; también se sugiere que algunas de las proteínas externas principales pueden tener una función de una porina2. Los patrones de proteínas de membrana externa parecen ser diversos entre las cepas de E. corrodens9; igualmente, se informó que estas proteínas inducen a bajas dosis la liberación de enzimas lisosomales a partir de macrófagos de ratones y exhiben efectos citotóxicos en dosis altas2,13; del mismo modo, presentan una estimulación dosis-dependiente o depresión de la fagocitosis y son capaces de disminuir la actividad del complemento y de inducir la agregación de plaquetas y trombina13. Los datos publicados hasta el momento sobre la función de las principales proteínas externas de membranas indican que pueden favorecer la actividad potencial de tipo patógeno en infecciones de E. corrodens, como en el desarrollo de las lesiones periodontales.
Complejo de adesina semejante a lectina. La adherencia bacteriana a las células huésped eucarióticas es dependiente de las interacciones entre una sustancia semejante a lectina y los receptores de superficie en las células de las mucosas humanas y/o de mamíferos. Para E. corrodens se informó la presencia de una proteína bacteriana parecida a lectina que media la adherencia a una molécula semejante a N-acetil-D-galactosamina14 en los receptores de superficie de las células humanas, como células epiteliales bucales, donde la actividad es sensible a tripsina o al calentamiento y se aumenta cuando se tratan estas células con neuraminidasa y a esta actividad la inhiben varios azucares que contienen D-galactosa y N-acetil-D-galactosamina14. La proteína semejante a lectina se ha purificado parcialmente, pues los análisis químico y electroforético, muestran que tiene posiblemente múltiples subunidades y forman un complejo proteínico14, que contiene hexosa pero no ácido murámico, glucosamina y ácido 2,6-diaminopimelico. Recientemente fue posible clonar, secuenciar y expresar un gen de E. corrodens que codifica un complejo parecido a lectina, con dos bandas de proteínas de peso molecular de 300 y 45 kDa en condiciones reductoras15; también, a la proteína de 45 kDa, componente del complejo y la principal proteína de la membrana externa, la codifica el gen porA que fue secuenciado y clonado y ofrece una alta homología con los genes de porinas de las especies de Neisseria, y puede jugar un papel importante en el proceso de adhesión bacteriana o de secreción de otros componentes con actividad semejante a lectina sobre la superficie celular15; finalmente, es importante mencionar el hallazgo de un gen que codifica una proteína de 25 kDa, que fue purificada y es un componente del complejo de adesina en E. corrodens16 y se encuentra cerca al dominio de reconocimiento de carbohidratos de la lectina en el complejo de la sustancia semejante a la lectina de E. corrodens. Los resultados en conjunto, señalan que la sustancia semejante a la lectina de E. corrodens es un factor importante en su adherencia a las células huésped, como una biopelícula17.
Pili. Los pilis son apéndices de superficie proteínicos, que funcionan como factores de virulencia para las especies de Neisseria y otras especies18, y la modulación de la piliación puede significar un mecanismo para evadir la respuesta inmune del huésped. La presencia de pili, otro factor de adhesión, se documentó bien por microscopía electrónica; están compuestos principalmente de una proteína denominada pilina tipo IV (Cuadro 3), que se ha clonado y secuenciado en dos tipos de genes de cepas diferentes de E. corrodens19. La pilina se purificó y caracterizó con un peso molecular aparente de 14.8 kDa y correlaciona la secuencia de aminoácidos del extremo N-terminal con otras pilinas de tipo IV19. En este mismo orden de ideas, E. corrodens exhibe una fase de variación irreversible que se refleja en los cambios de la morfología de colonia, observándose colonias pequeñas (S) con morfología corrodens y grandes (L) con morfología no corrodens; esta variación de fase se relaciona con la presencia de pili en las pequeñas y su ausencia en las grandes; del mismo modo, se ha tratado de explicar desde el punto de vista molecular esta variación de fase, presente en otros gérmenes patógenos como N. gonorrhoeae, donde las variantes L sintetizan la pilina pero no la exportan y ensamblan en pili, mientras que las de la fase S la exportan y ensamblan en pili, debido posiblemente a un evento postranslacional20.
La estructura y función del pili se analizó en una cepa clínica de E. corrodens VA1, donde se definió el locus pilA que incluye pilA1, pilA2, pilB y hagA20. El pilA1 codifica la proteína pili de mayor peso molecular de esta cepa; pero, la inactivación de pilA1 produjo una mutante que fue fenotípicamente indistinguible de las variantes de fase-L, lo que indicó que la biosíntesis del pili tipo IV depende de la expresión del pilA1 y de la apropiada exportación y ensamblaje de pilA1. Otros experimentos con mutantes inactivas para pilA2, pilB y hagA, mostraron que sólo aquellas inactivadas para pilB fueron deficientes para motilidad de contracción (twitching), lo que sugiere una función para pilB en este fenómeno20.
En resumen, los datos muestran que E. corrodens sintetiza una proteína pilina tipo IV que se encuentra en el locus pilA y presenta una transición irreversible de colonias pequeñas a grandes; finalmente, se postula que la relación de los procesos de adhesión y la modulación en la formación de pili (variación antigénica), puede representar un mecanismo para evadir la respuesta inmune del huésped.
Otros mecanismos de patogénesis. Los mecanismos de patogénesis de E. corrodens se relacionan con sus características morfológicas de colonia, actividades enzimáticas, factores de agregación e interacciones con otras especies y diversidad clonal (Cuadro 4); asimismo, es un factor básico en el proceso de infecciones la colonización de los tejidos por parte del microorganismo. Los estudios sobre las morfologías de las colonias de E. corrodens mediante microscopía electrónica han encontrado que la translocación de superficie denominada “movilidad de contracción”o “motilidad de deslizamiento” ocurre en la porción externa de la colonia; pero, es posible que esta motilidad y la variación en la morfología de la colonia (corrodens o no corrodens, pequeñas o grandes) cumplan algún papel en el proceso de colonización en las áreas periodontales2,11
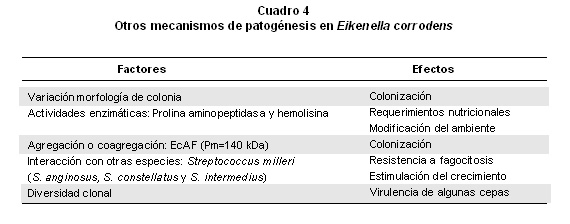
E. corrodens presenta actividades de prolina aminopeptidasa y hemolisina dependiente de tiol que pueden ser centrales para su patogenicidad3, 4, 21. La hemolisina puede poseer actividad de esterasa, pues no se ubica dentro de la categoría de fosfolipasa de hemolisinas, que dañan la membrana porque no tiene actividad contra lecitina y no se encuentra en el grupo de citolisinas activadas por tiol porque el colesterol no la inhibe3. Por otro lado, la actividad de prolina iminopeptidasa puede estar relacionada con los requerimientos nutricionales de E. corrodens porque necesita modificar las condiciones del ambiente local al igual que protegerse contra los mecanismos de defensa del huésped, siendo significativos el colágeno, la región de bisagra de las inmunoglobulinas y proteínas del complemento que son ricas en prolina y pueden actuar posiblemente como substratos3. También, se han descrito otras actividades sobre glutamato, serina y glutamina que resultan en altos niveles de reducción de nitrato4, lo cual le sería ventajoso por las características del medio en los sitios periodontalmente enfermos; asimismo, la importancia de la actividad de prolina arilamidasa que puede ser útil como marcador diagnóstico para identificar este microbio patógeno21.
Las bacterias pueden agregarse en forma de grupos (clumps) dentro de la misma especie o coagregarse en una comunidad compleja de biopelícula17; a estos procesos los pueden mediar anticuerpos específicos de la saliva y factores glucoproteínicos no específicos. La agregación o coagregación pueden ser parte del proceso de colonización o de defensa del huésped; pero, cuando los grupos llegan a ser suficientemente grandes, serían susceptibles de limpieza mecánica por fuerzas de cizalla o flujo salival21. Para E. corrodens se ha descrito un factor de agregación salival glucoproteínico (EcAF) con un peso molecular aparente de 140 kDa, inestable frente al calor o al tratamiento con proteasa e inhibido por N-acetil-D-galactosamina y sacáridos que contienen un residuo de galactosa en el extremo reducido; además, la actividad de agregación fue sensible a EDTA y restablecida por Ca+2, pero no por Mn+2 o Mg+222; el tratamiento con neuraminidasa de EcAF aumentó la capacidad de agregación de E. corrodens, e indica que los ácidos siálicos sobre EcAF interfieren con la agregación.
E. corrodens suele encontrarse junto con otros microorganismos en los sitios donde se ha aislado, por lo que es probable que actúe como parte de un consorcio microbiano predominante2-4,23, lo cual se refleja en que se puede asociar con Porphyromonas gingivalis, Prevotella intermedia, Actinobacillus actinomycetemcomitans, Capnocytophaga spp., Campylobacter rectus y Fusobacterium nucleatum. Se han descrito interacciones con el grupo de Streptococcus milleri (S. intermedius, S. contellatus y S. anginosus) en las que la co-agregación y la estimulación del crecimiento que se presentan entre estas especies pueden ser relevantes para establecer infecciones mixtas que comprometen a estas bacterias23, porque se ha demostrado que las células coagregadas son más resistentes a fagocitosis y a la muerte por neutrófilos tanto in vitro como in vivo que los microorganismos solos, lo que explica la patogénesis de las infecciones extra-orales, debido a que estos agregados de placa bacteriana entran al torrente sanguíneo y causan una infección focalizada desde la cavidad oral23; asimismo, el metabolismo de las especies de Streptococcus lo puede influir E. corrodens pues sus nichos metabólicos pueden coincidir y aprovechar sus actividades hidrolíticas de manera conjunta sobre los sustratos disponibles, que pueden reflejar el aumento de la actividad enzimática o la producción de un factor estimulante del crecimiento.
Puede existir una relación estrecha entre las enfermedades cardíacas y la enfermedad periodontal, pues hay una vía de activación inflamatoria mediada por LPS y monocitos24,25. En la enfermedad coronaria y en la aterosclerosis humanas intervienen el factor-a de necrosis tumoral (TNF-a), 1b-interleuquina (IL-1b) y prostaglandina E2, PGE2. Los LPS bacterianos inician la expresión de IL-a que impide la fibrinólisis, pero agiliza la coagulación y la trombosis; mientras, el TNF-a y la IL-1b elevan la acumulación de colesterol en los monocitos y la irradiación del músculo liso, lo cual origina paredes vasculares más gruesas24. Por otro lado, en la enfermedad periodontal, los monocitos responden a los LPS de la placa bacteriana por secreción de mediadores pro-inflamatorios como TNF-a, IL-1b y PGE2 y tromboxano A2(TxA2), que no sólo dañan el periodonto sino que inducen una respuesta sistémica a la penetración de la bacteria en los tejidos, pérdida de la integridad del epitelio en la bolsa periodontal y/o bacteriemia transitoria.
Se ha demostrado que la endotoxina de los microorganismos de la placa es capaz de penetrar el espacio gingival y tiene cantidades suficientes de LPS para lograr una respuesta sistémica y específica de anticuerpos LPS25, lo que indica la presencia de cantidades apreciables de LPS para gérmenes periodontopatógenos; de esta manera, las citoquinas y los LPS del periodonto infectado que entran a la circulación sistémica serían los responsables de iniciar o empeorar las condiciones asociadas con aterosclerosis y enfermedad coronaria24,25. Si bien es cierto que a E. corrodens se le asocia con enfermedad coronaria, también en los estudios in vitro se ha encontrado que la cepa silvestre de esta especie prácticamente carece de la capacidad para invadir células endoteliales y células de músculo liso de arterias coronarias humanas si se compara con otras especies como P. gingivalis, tal vez porque ciertas células fagocitan a E. corrodens y así se evita una invasión activa por esta bacteria26; sin embargo, la eficiencia en la invasión de las células como se observa en P. gingivalis es variable entre cepas diferentes, pues se ha visto que falta tal habilidad en una cepa de esta especie26; de igual manera, la presencia de adesinas no es suficiente para la invasión, porque si una bacteria no puede adherirse, no puede invadir. Estos hallazgos quizá no sean concluyentes para sustentar el posible papel de E. corrodens en las enfermedades cardiovasculares, pero sí permitirían establecer una relación indirecta entre E. corrodens y enfermedad cardiovascular y a estab-proteobacteria como un agente causal de enfermedad periodontal en asocio con otros microorganismos.
Otro factor relacionado con la patogenicidad de la bacteria es la característica de la diversidad clonal de E. corrodens11,27-30. Por análisis de endonucleasas de restricción se encontraron en un enfermo seis clones distintos y dos tipos diferentes en una sola bolsa periodontal27. Por el método de reacción en cadena de la polimerasa (PCR) se ha descubierto en el proceso de colonización la presencia de múltiples clones en la cavidad oral y se sugiere una complejidad en la flora microbiana gingival28. La presencia de clones distintos genéticamente en el mismo sujeto puede ser muy significativa para comprender la patogenicidad de E. corrodens pues algunas cepas pueden ser virulentas y capaces de causar infección, mientras otras pueden ser relativamente inocuas y constituir un componente normal en la flora de la boca; además, se informó que el plásmido pMU1 puede estar comprometido en la patogenicidad y virulencia de E. corrodens31. Igualmente, esta diversidad puede influir en la respuesta inmunológica del huésped, como se ha observado en los niveles variables de inmunoglobulina G sérica, lo cual es importante para evaluar la respuesta inmune frente a este microorganismo29. Sobre la respuesta inmune inducida por E. corrodens, experimentalmente se ha descrito el estímulo en las secreciones de IL-6 y IL-8 cuando está en contacto directo con células epiteliales de la boca, aunque de necesidad no se requiere para la respuesta, lo cual sugiere que la secreción de citoquinas pro-inflamatorias puede servir como un signo inicial para el sistema inmune del huésped30.
INFECCIONES ORALES Y NO ORALES
E. corrodens es un habitante normal de la cavidad oral y del tracto respiratorio superior, y las evidencias sugieren que es un microorganismo oportunista que en general se asocia con otros gérmenes en enfermedades orales y no orales. Chen y Wilson2 documentaron bien la relación entre E. corrodens y enfermedad periodontal e infecciones no orales, mediante estudios en animales y estudios epidemiológicos longitudinales y de corte o transversales, los cuales sugieren que:
E. corrodens es un microorganismo particularmente virulento en animales de laboratorio. Como organismo patógeno poco frecuente, exhibe un comportamiento oportunista, pues casi todas las infecciones no orales se consideran como resultado de la transmisión del germen desde la cavidad oral humana.
En la flora normal de la boca es un habitante común, por tanto la presencia de E. corrodens en la placa subgingival es compatible con periodonto sano.
Los resultados de los estudios transversales no son concluyentes, pero muestran que una apreciable cantidad de E. corrodens se halla en los sitios enfermos; en consecuencia, puede participar de alguna manera en la progresión de la destrucción periodontal.
Los estudios longitudinales sugieren un papel limitado en la patogénesis de la enfermedad periodontal.
FACTORES PREDISPONENTES
En estudios recientes se han descrito factores que predisponen a infecciones por la presencia de E. corrodens (Cuadro 5) en individuos con periodontitis y hábito de fumar, VIH seropositivos, daño genético, síndrome de Down, y síndrome de Papillon-Lefèvre y la enfermedad de Behçet’s. Los sitios de infección extra oral descritos hasta ahora para E. corrodens son diversos; por ejemplo, cabeza y cuello, extremidades, abdomen, sangre, válvulas cardíacas, sistema nervioso central, aparato genital, ojos, glándula tiroides, tejido óseo; pero, los casos en mayor proporción se presentan en las extremidades, principalmente con las mordeduras humanas2. En un estudio reciente durante un período de 6 años (1993-1998) en Taiwán se analizaron 43 casos de infecciones invasivas por diversos clones de E. corrodens32, con resultados que indican una mayor proporción de casos de infección en cabeza y cuello (56%) e infección pulmonar (23%), y en menor porcentaje casos de infección intra-abdominal (14%), infección cutánea (5%), infección ósea (2%), endocarditis (2%) y absceso pélvico (2%); además, 63% presentaban enfermedades de base, sobre todo malignidad en cabeza y cuello en 35% y 56%, respectivamente, presentaron factores que predisponían la invasión. Las infecciones polimicrobianas sucedieron en 65% de los pacientes y se aislaron estreptococos en 66%; hubo, cinco casos fatales, con cuatro fallecimientos atribuibles directamente a la propagación de la infección de E. corrodens.
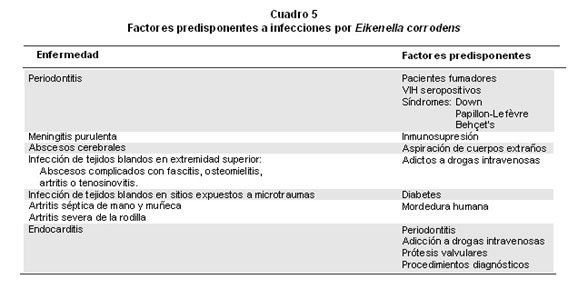
E. corrodens se puede presentar como agente etiológico en meningitis purulenta asociada con inmunosupresión e infecciones polimicrobianas de tejidos blandos en sujetos adictos a drogas intravenosas33. En un estudio retrospectivo de 57 drogadictos se encontraron abscesos en la extremidad superior34 que se complicaron con fascitis, osteomielitis, artritis o tenosinovitis séptica y nueve resultaron VIH sero-positivos; se hallaron como bacterias más comunes Streptococcus, Staphylococcus aureus y E. corrodens. De igual forma, E. corrodens aparece como agente causal de artritis séptica de la mano y la muñeca, artritis severa de la rodilla, provocada por lo general por traumas como mordedura animal o humana35; y también, en un caso de úlcera del pene asociada con sexo oral (fellatio)36.
Quienes padecen diabetes sufren de micro traumas en la piel, producto del monitoreo diario de la glucosa y las inyecciones de insulina, pues se exponen a la infección por E. corrodens a través de secreciones orales cuando éstas entran en contacto con los traumas de la piel. Se informaron dos casos de infecciones por E. corrodens en dos jovencitas con diabetes37, una tenía cuatro años de evolución y estaba moderadamente controlada; en ella las punciones rutinarias en un dedo indujeron un panadizo complicado sin respuesta a la administración de cefalexina oral, y produjo necrosis de la yema del dedo y del lecho de la uña. La segunda, con evolución de cinco años y control muy pobre desarrolló un absceso agudo en el muslo, sitio de inyección de la insulina, que se resolvió después de drenaje y antibióticos intravenosos. Finalmente, E. corrodens se ha relacionado con endocarditis infecciosa38 (Cuadro 5) en asocio no sólo con enfermedad periodontal de base39 y el uso de drogas intravenosas33 sino con procesos diagnósticos como colonoscopia y biopsia de mucosa gástrica40; además, con otras entidades como linfadenopatía inmunoblástica donde se une con otros microorganismos como Streptococcus del grupo C41. Se le ha aislado también con especies de Actinomyces en pacientes de osteomielitis crónica difusa esclerosante42 que ocurre sólo en la mandíbula de mujeres jóvenes y se caracteriza por ser muy dolorosa.
ENDOCARDITIS
La endocarditis infecciosa es la más grave de las infecciones endovasculares, producida por microorganismos oportunistas encontrada en la práctica clínica. E. corrodens se presenta como agente causal de endocarditis de manera rara, por lo general en personas con prótesis valvulares40,43 y adictos a drogas intravenosas33. Se describe como parte del grupo HACEK (Haemophilus parainfluenzae, H. aphrophilus y H. paraphrophilus, Actinobacillus actinomycetemcomitans, Cardiobacterium hominis, Eikenella corrodens y Kingella kingae) que son agentes más raros en la etiología de endocarditis infecciosa38,39; asimismo, otros organismos, como hongos, son responsables hasta de 15% de los casos de endocarditis infecciosa de prótesis valvulares39; mientras, los microorganismos del grupo HACEK causan 3% de casos de endocarditis infecciosa adquirida en la comunidad. En pacientes con endocarditis y una historia de enfermedad periodontal o trabajo dental en quienes los cultivos de sangre rutinarios son negativos podrían tener infección debida a variantes de estreptococos o bacterias del grupo HACEK38,39 por efecto del factor nutricional.
Con respecto a pacientes con prótesis valvulares se han informado casos por E. corrodens con enfermedad prolongada debido a infecciones de prótesis vasculares periféricas. Cuando se presenta como agente único, las infecciones de los espacios vasculares tienden a ser indoloras, pero puede ser necesario retirar las prótesis vasculares43. Casi siempre las infecciones por E. corrodens derivan de la cavidad oral, pero se supo de un caso donde lo más probable es que se haya adquirido por biopsia de mucosa en una colonoscopia, que se hizo al paciente pocos días antes de aparecer la enfermedad40. En otro individuo adicto a drogas intravenosas se encontró vegetación tricúspide pedunculada, ventrículo derecho dilatado, disminución de la contractilidad y regurgitación.
OSTEOMIELITIS
E. corrodens se vio en osteomielitis o infecciones de cabeza y cuello, ya sea como agente causal primario o en asocio con otras bacterias42,44, en drogadictos como una complicación en casos de abscesos en las extremidades superiores34; también, se aisló en pacientes con mordeduras humanas en la mano, donde además puede haber artritis y celulitis45. Stoloff & Gillies46 informaron que de 33 personas con infecciones por E. corrodens en un hospital general durante un período de 18 meses, 18 pacientes se relacionaban con este tipo de trauma y en nueve hubo complicación por osteomielitis y/o artritis séptica.
INFECCIONES INTRA-ABDOMINALES
En las infecciones intra-abdominales se ha descrito a E. corrodens como agente etiológico único o en asocio con otros microorganismos; además, estas infecciones son usualmente indoloras y la formación de abscesos se da universalmente. Gonzálvez et al.47 revisaron los resultados del laboratorio bacteriológico del Hospital General de Albacete (España) por un período de cinco años, y encontraron que E. corrodens se aisló en niños de 1 a 7 años con peritonitis secundaria a apendicitis perforada; al mismo tiempo, hasta la fecha del estudio se diagnosticaron 30 casos de infecciones intra-abdominales por E. corrodens de los cuales 17 eran en niños. La revisión de Danziger et al.48 reveló 19 casos de infección intra-abdominal, donde 11 pacientes eran menores de 25 de años de edad, y se aisló E. corrodens junto con otras bacterias, sobre todo varias especies de estreptococos. También, se informó como agente causal de abscesos hepáticos de carácter polimicrobiano, como resultado probable de la extensión de infecciones orales o abdominales49.
INFECCIONES DEL SISTEMA NERVIOSO CENTRAL
Emmerson & Mills50 encontraron E. corrodens en un enfermo de otitis media previa y mastoidectomía como causa de absceso cerebral y de meningitis recurrente. Pit et al.51, en Malasia, hallaron 75 casos de absceso cerebral en los que había como condiciones predisponentes la otitis media supurativa crónica, enfermedad cardíaca congénita y meningitis. Los cultivos puros, 66.7%, predominaron y los estériles se obtuvieron en 13.3% de los casos; además, se aislaron estreptococos en 23 pacientes, donde la especie más común fue S. milleri; pero, en cultivos puros se encontró E. corrodens en sujetos con enfermedad cardíaca congénita cianótica51.
DIAGNÓSTICO
Para el diagnóstico de estas enfermedades se deben evaluar las características de E. corrodens con respecto al crecimiento en agar sangre y/o chocolate como colonias que corroen el agar y su motilidad de contracción, así como las pruebas bioquímicas y otras que permitan su correcta identificación (Cuadro 6). Se han descrito medios de cultivo selectivos para E. corrodens a partir de lesiones periodontales52 y otros medios que permiten el crecimiento de bacterias fastidiosas como E. corrodens53. El análisis de los datos para identificar E. corrodens permite recomendar, que el material clínico requiere un tratamiento previo con el objetivo de homogeneizar la muestra biológica, y el enriquecimiento se realiza en un medio sólido; por ejemplo, agar de infusión cerebro, corazón, tripticasa de soya, Todd Hewitt y chocolate suplementado con extracto de levadura, nitrato de potasio, hemina y sangre de cordero; además, se le deben adicionar 25 mg de clindamicina/ml. El agar seleccionado con el suplemento se puede incubar en condiciones anaeróbicas o microaerofílicas a temperatura de 34ºC por 48 a 72 horas. En este tiempo se observan colonias que corroen y piquetean el agar y la identificación como E. corrodens se confirma por criterios morfológicos y bioquímicos1,3-5. Por otro lado, la cuantificación de la actividad de prolina iminopeptidasa con L-prolina-p-nitroanilida como substrato3,21 se ha propuesto como indicador de E. corrodens en la infección periodontal; asimismo, se le puede detectar si se evalúa la respuesta inmunológica29 por la prueba de inmuno absorción enzimática (ELISA). El uso de otras técnicas como PCR y sondas de ADN para su identificación podrían ser simples y rápidas en muestras clínicas especialmente periodontales27,28,54.

TRATAMIENTO
Se conoce la resistencia de E. corrodens a clindamicina, metronidazol, cefalotina, vancomicina, cefuroxima32,55. Sin embargo, los antimicrobianos que tienen una estructura derivada del anillo quinolona y/o fluoroquinolona56-59 fueron activos contra 16 a 151 cepas de E. corrodens aisladas de muestras clínicas (Cuadro 7), con una MIC50 y MIC90 entre 0.008-0.12 y 0.015-0.125 mg/ml, respectivamente; mientras, los antibióticos con una estructura penem60,61 muestran una actividad en un rango de 0.015 a 0.25 mg/ml; pero los agentes ketólidos62,63, macrólidos sintéticos derivados de la eritromicina que inhiben la biosíntesis de las proteínas, la ampicilina y minociclina tienen una MIC90 de 1.0 a 2.0 mg/ml64; además, las MICs de las cefalosporinas cefotetan, cefotaxima, ceftrioxone y cefepime56,60,62-64 que inhibieron 50% y 90% de las cepas aisladas fueron 0.015-0.5 y 0.015-1.0 mg/ml, respectivamente. Para aumentar la sensibilidad puede utilizarse la combinación de dos antibióticos56,57,60-64 donde uno de ellos es el clavulanato o sulbactam o tazobactan (Cuadro 7). E. corrodens presenta una sensibilidad mediana a los antimicrobianos que tienen una MIC90 entre 2.0 y 4.0 mg/ml (Cuadro 7); por ejemplo, doxiciclina, tetracilina, cefoxitina, eritromicina, cefuroxime, etc.
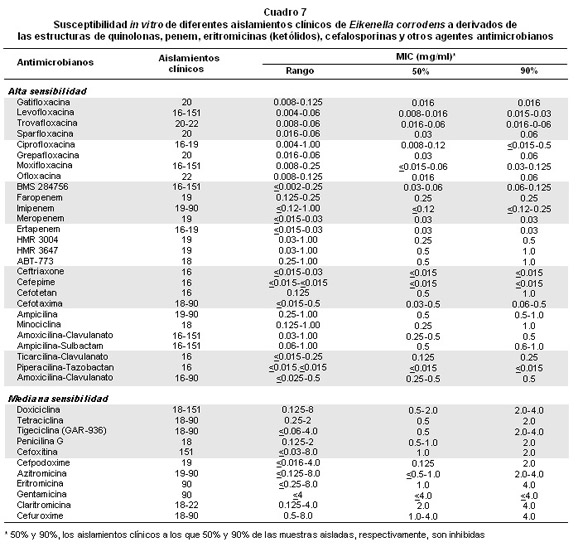
El análisis de los resultados anteriores sugiere que los antibióticos de alta sensibilidad tienen un papel terapéutico potencial en el tratamiento de las infecciones por E. corrodens sola y/o en consorcio microbiano. Pero, es importante destacar que como primera opción contra la actividad in vitro de este germen patógeno, se encuentran los antimicrobianos que tienen estructuras derivadas de la quinolona; de la misma forma, como segunda elección se hallan los antibióticos con estructuras penem; finalmente, las cefalosporinas cefepime, ceftrioxone y cefotaxima, como también la combinación de los antibióticos piperacilina-tazobactan y ticarcilina-clavulanato. Además, esta actividad in vitro requiere una evaluación como alternativa terapéutica en infecciones humanas.
PERSPECTIVAS
E. corrodens es un microbio patógeno en infecciones orales y no orales, que tiene un carácter oportunista y de manera general actúa en asocio con otras especies, que se relaciona con diversas entidades que tienen en común la presencia de ciertos factores de riesgo donde puede ser difícil la asociación con este microorganismo. Se le ha aislado en pacientes con periodontitis, parotiditis, sinusitis, osteomielitis, absceso oral y no oral, bacteriemia, endocarditis, infecciones intra-abdominales, del sistema nervioso central y pulmonares1,2,23,24,29,38; además, los resultados indican la presencia de especies con genotipos y fenotipos diferentes de E. corrodens28,31, aun en un mismo paciente y puede ser muy significativo para comprender la patogenicidad en los seres humanos cuando se encuentra solo y/o en consorcio microbiano en el sitio de infección.
Los resultados señalan que los factores de virulencia se asocian con los polisacáridos, limo, proteínas de membrana externa, complejo de adesina semejante a lectina, pili, característica de colonia, actividades enzimáticas y diversidad clonal. Sin embargo, es importante estudiar con mayor profundidad, desde el punto de vista molecular la composición, funciones, características y relaciones entre la(s) molécula(s) de los distintos factores de virulencia y el sitio de infección, de tal manera que permita una visión más completa de la interacción de E. corrodens y el huésped. Asimismo, se ha demostrado que en diferentes nichos ecológicos E. corrodens hace parte de un consorcio microbiano, pero se requiere identificar con mayor detalle la naturaleza de la coagregación y las reacciones que estimulan el crecimiento, como el empleo de algunos productos del metabolismo de los microorganismos presentes en la infección. También, se debe ampliar la investigación de la actividad proteolítica y la composición del medio de cultivo que permita un amplio crecimiento de E. corrodens, así como su metabolismo en sentido catabólico y en los procesos anabólicos, en especial los fenómenos de transporte y los aspectos bioenergéticos. Aunque se ha avanzado en la investigación biomédica y básica3,5,65, es indispensable que las futuras investigaciones sobre la bioenergética de E. corrodens deban dirigirse a profundizar la presencia de otros sustratos fisiológicos y los citocromos de la cadena respiratoria de E. corrodens5, y seguir en los estudios sobre el efecto de inhibidores y análogos de quinonas en el flujo de electrones; igualmente, iniciar el análisis del sistema transportador de electrones en condiciones anaeróbicas; además, es importante la purificación y caracterización de los diversos complejos respiratorios, que suministraría información sobre la función de cada uno de ellos y funcionamiento del sistema respiratorio en diferentes tensiones de oxígeno y relacionarla con los mecanismos de patogenicidad in vivo. Por último, la disponibilidad de las secuencias del genoma para Neisseria sp., podría facilitar el diseño de vectores genómicos para clonar genes respiratorios y virulentos similares de E. corrodens.
La caracterización de un microorganismo fastidioso como E. corrodens, que no es fácil aislar y diagnosticar, por tanto representa un desafío para el clínico, por lo que es importante generar conocimiento biomédico y básico, así como estudiar aún más el crecimiento selectivo en agar especial, pruebas bioquímicas, características serológicas, ensayos en la región visible del espectrofotómetro, estrategias de histoquímica, ELISA, PCR, sondas de ADN y microscopía electrónica, para permitir un rápido y oportuno tratamiento al paciente. La variable susceptibilidad a los antimicrobianos por las distintas muestras clínicas aisladas de E. corrodens, sugiere realizar estudios con moléculas altamente específicas para inhibir la replicación y/o transcripción y/o síntesis de proteínas y/o el funcionamiento de la cadena respiratoria en el complejo bc1 la respiración y/o citocromo oxidasa y/o nitrato reductasa, a fin de obtener un fármaco y/o combinaciones diversas con más alto potencial terapéutico en el tratamiento de infecciones en que se presente estab-proteobacteria sola o en consorcio microbiano.
REFERENCIAS
1. Jackson FL, Goodman YE. Transfer of the facultatively anaerobic organism Bacteroides corrodens Eiken, to a new genus, Eikenella. Int J Syst Bacteriol 1972; 22: 73-77.
2. Chen C KC, Wilson ME. Eikenella corrodens in human oral and non-oral infections: A review. J Periodontol 1992; 63: 941-953.
3. Allaker RP, Young KA, Hardie JM. Production of hydrolytic enzymes by oral isolates of Eikenella corrodens. FEMS Microbiol Letts 1994; 123: 69-74.
4. Gully NJ, Rogers AH. Energy production and peptidase activity in Eikenella corrodens. FEMS Microbiol Letts 1996; 139: 209-213.
5. Jaramillo RD, Barraza BC, Polo A, Sará M, Contreras M, Escamilla JE. The aerobic electron transport system of Eikenella corrodens. Can J Mcrobiol 2002; 48: 895-902.
6. Progulske A, Holt SC. Isolation and characterization of the outer membrane and lipopolysaccharide from Eikenella corrodens. Infect Immun 1984; 43: 166-177.
7. Progulske A, Mishell R, Trummel C, Holt SC. Biological activities of Eikenella corrodens outer membrane and lipopolysaccharide. Infect Immun 1984; 43: 178-182.
8. Kato T, Takasoe I, Okuda K. Structural analysis of lipopolysaccharides from Eikenella corrodens by use of murine monoclonal antibodies. Infect Immun 1989; 57: 656-659.
9. Chen C KC, Wilson ME. Outer membrane protein and lipopolysaccharide heterogeneity among Eikenella corrodens isolates. J Infect Dis 1990; 162: 664-671.
10. Shiozu I, Onagawa M, Okuda K. A heat stable hemagglutinating activity of Eikenella corrodens. Bull Tokio Dent Coll 1991; 32: 57-61.
11. Coob CM, Helber JT, Hirschberg R. Scanning electron microscopy of Eikenella corrodens colony morphology variants. J Periodont Res 1994; 29: 410-417.
12. Behling U H, Pham P H, Nowotony A. Biological activity of the slime and endotoxin of the periodontopathic organism Eikenella corrodens. Infect Immun 1979; 26: 580-584.
13. Tufano MA, Sommese L, Galdiero F. Some biological activities of Eikenella corrodens major outer membrane proteins. Eur J Epidemiol 1986; 2: 305-311.
14. Yamasaki Y, Ebisu S, Okada H. Partial purification of bacterial lectin-like substance from Eikenella corrodens. Infect Immun 1988; 56: 191-196.
15. Yumoto H, Azakami H, Nakae H, Matsuo T, Ebisu S. Cloning, sequencing and expression of an Eikenella corrodens gene encoding a component protein of the lectin-like adhesin complex. Gene 1996; 183: 115-121.
16. Azakami H, Yumoto H, Nakae H, Matsuo T, Ebisu S. Molecular analysis of the gene encoding a protein component of the Eikenella corrodens adhesin complex that is close to the carbohydrate recognition domain. Gene 1996; 180: 207-212.
17. Azakami H, Nakashima H, Akimichi H, Noiri Y, Ebisu S, Kato A. Involvement of N-Acetyl-D-galactosamine-specific lectin in biofilm formation by the periodonto-pathogenesis bacterium, Eikenella corrodens. Biosci Biotechnol Biochem 2006; 70: 441-446.
18. Aho EL, Keating AM, McGillivray, A. A comparative analysis of pilin from pathogenesis and
nonpathogenesis Neisseria species.
Microb Pathog 2000; 28: 81-88.
19. Hood BL, Hirschberg R. Purification and characterization of Eikenella corrodens type IV pilin. Infect Immun 1995; 63: 3693-3696.
20. Villar MT, Hirschberg RL, Schaefer MR. Role of the Eikenella corrodens pilA locus in pilus function and phase variation. J Bacteriol 2001; 183: 55-62.
21. Allaker RP, Young KA, Hardie JM. Rapid detection of proline iminopeptidase as an indicator of Eikenella corrodens periodontal infection. Lett Appl Microbiol 1994; 19: 325-327.
22. Ebisu S, Nakae H, Fukuhara H, Okada H. The mechanisms of Eikenella corrodens aggregation by salivary glycoprotein and the effect of the glycoprotein on oral bacterial aggregation. J Periodont Res 1992; 27: 615-622.
23. Young KA, Allaker RP, Hardie JM, Whiley RA. Interactions between Eikenella corrodens and Streptococcus milleri-group organisms: possible mechanisms of pathogenicity in mixed infections. Antonie Van Leeuwenhoek 1996; 69: 371-373.
24. Meyer DH, Fives-Taylor PM. Oral pathogens: from dental plaque to cardiac disease. Curr Opin Microbiol 1998; 1: 88-95.
25. Kinane DF, Lowe G DO. How periodontal disease may contribute to cardiovascular disease. Periodontol 2000; 23: 121-126.
26. Dorn BR, Dunn WA, Progulske-Fox A. Invasion of human coronary artery cells by periodontal pathogens. Infect Immun 1999; 67: 5792-5798.
27. Chen C, Sunday GJ, Zambon JJ, Wilson ME. Restriction endonuclease analysis of Eikenella corrodens. J Clin Microbiol 1990; 28: 1265-1270.
28. Fugise O, Chen W, Rich S, Chen C. Clonal diversity and stability of subgingival Eikenella corrodens. J Clin Microbiol 2004; 42: 2036-2042.
29. Chen C, Ashimoto A. Variable serum immunoglobulin G immune response to genetically distinct Eikenella corrodens strains coexisting in the human oral cavity. Oral Microbiol Immunol 1998; 13: 158-165.
30. YumotoH, Nakae H, Fujinaka K, Ebisu S, Matsuo T. Interleukin-6 (IL-6) and IL-8 are induced in human oral epithelial cells in response to exposure to periodontophatic Eikenella corrodens. Infect Immun 1999; 67: 384-394.
31. Azakami H, Akimichi H, Noiri Y, Ebisu S, Kato A. Plasmid-mediated genomic recombination at the pilin gene locus enhances the N-acetyl-D-galactosamine-specific haemagglutination activity and the growth rate of Eikenella corrodens. Microbiol 2006; 152: 815-821.
32. Sheng WS, Hsueh PR, Hung CC, Teng LJ, Chen YC, Luh KT. Clinical features of patients with invasive Eikenella corrodens infections and microbiological characteristics of the clinical isolates. Eur J Clin Microbiol Infect Dis 2001; 20: 231-236.
33. Olopoenia LA, Mody V, Reynolds M. Eikenella corrodens endocarditis in an intravenous drug user: case report and literature review. J Natl Med Assoc 1994; 86: 313-315.
34. Gonzáles MH, Garst J, Nourbash P, Pulvirenti J, Hall RF. Abscesses of the upper extremity from drug abuse by injection. J Hand Surg (Am) 1993; 18: 868-870.
35. Murray P. Septic arthritis of the hand and wrist. Hand Clin 1998; 14: 579-587.
36. Rosen T. Penile ulcer from traumatic orogenital contact. Dermatol Online J 2005; 11: 18.
37. Newfield RS, Vargas I, Huma Z. Eikenella corrodens infections. Case report in two adolescent females with IDDM. Diabetes Care 1996; 19: 1011-1013.
38. Cardoso KN, Ochiai ME, Oliveira MT, Morgado P, Munhoz R, Andretto FE, Mansur AJ, Barreto AC. Eikenella corrodens infective endocarditis. Arq Bras Cardiol 2005; 85: 63-64.
39. Berbari EF, Cockerill FRR, Steckelberg JM. Infective endocarditis due to unusual or fastidious microorganisms. Mayo Clin Proc 1997; 72: 532-542.
40. Heiro M, Nikoskelainen J, Engblom E, Kotilainen P. Eikenella corrodens prosthetic valve endocarditis in a patient with ulcerative colitis. Scand J Infect Dis 2000; 32: 324-325.
41. Mönkemüller KE, Bronze MS. Immunoblastic lymphadenopathy presenting as an acute abdomen and mixed bacteriemia with Eikenella corrodens and group C streptococci. Am J Gastroenterol 1998; 93: 652-653.
42. Marx RE, Carlson ER, Smith BR, Toraya N. Isolation of Actinomyces species and Eikenella corrodens from patients with chronic diffuse sclerosing osteomyelitis. J Oral Maxillofac Surg 1994; 52: 26-33.
43. Decker MD, Graham BS, Hunter EB, Liebowitz SM. Endocarditis and infections of intravascular devices due to Eikenella corrodens. Am J Med Sci 1986; 292: 209-212.
44. Jones JL, Romig DA. Eikenella corrodens: a pathogen in head and neck infections. Oral Surg Oral Med Oral Pathol Oral Radiol 1979; 48: 501-505.
45. Johnson SM, Pankey GA. Eikenella corrodens osteomyelitis, arthritis, and cellulitis of the hand. South Med 1976; 69: 535-539.
46. Stoloff AL, Gillies ML. Infections with Eikenella corrodens in a general hospital: a report of 33 cases. Rev Infect Dis 1986; 8: 50-53.
47. Gonzálvez PJ, Marco MA, Pérez MA. Intraabdominal infections caused by Eikenella corrodens in children. Cir Pediatr 1995; 8: 145-147.
48. Danziger LH, Schoonover LL, Kale P, Resnick DJ. Eikenella corrodens as an intra abdominal pathogen. Am Surg 1994; 60: 296-299.
49. Chang PS, Ni YH, Lin WT, Lee CY, Chang MH. Isolation of Eikenella corrodens from polymicrobial hepatic abscess: report of one case. Acta Paediatr Taiwan 1999; 40: 50-52.
50. Emmerson AM, Mills F. Recurrent meningitis and brain abscess caused by Eikenella corrodens. Postgrad Med J 1978; 54: 343-345.
51. Pit S, Jamal F, Cheah FK. Microbiology of cerebral abscess: a four-year study in Malaysia. J Trop Med Hyg 1993; 96: 191-196.
52. Goldstein EJC, Agyare EO, Silleti R. Comparative growth of Eikenella corrodens on 15 media in three atmospheres of incubation. J Clin Microbiol 1981; 13: 951-953.
53. Scythes KD, Louie M, Simor AE. Evaluation of nutritive capacities of 10 broth media. J Clin Microbiol 1996; 34: 1804-1807.
54. Furcht C, Eschrich K, Merte K. Detection of Eikenella corrodens and Actinobacillus actinomycetemcomitans by use of the polymerase chain reaction (PCR) in vitro and in subgingival plaque. J Clin Periodontol 1996; 23: 891-897.
55. Aracil B, Alós JI, Gómez Garcés JL. Eikenella corrodens y antimicrobianos orales, clásicos y recientes. Rev Esp Quimioter 1997; 10: 156-160.
56. Goldstein EJC, Citron DM, Merriam CV, Warren Y, Tyrrel K, Fernandez H. In vitro activities of the Des-fluro(6) quinolone BMS-284756 against aerobic and anaerobic pathogens isolated from skin and soft-tissue animal and human bite wound infections. Antimicrob Agents Chemother 2002; 46: 866-870.
57. Goldstein EJC, Citron DM, Merriam CV, Warren Y, Tyrrel K, Fernandez H. In vitro activities of a new Des-fluroquinolone BMS-284756, and seven other antimicrobial agents against 151 isolates of Eikenella corrodens. Antimicrob Agents Chemother 2002; 46: 1141-1143.
58. Goldstein EJ, Citron DM, Merriam CV, Tyrrell K, Warren Y. Activity of gatifloxacin compound to those of five other quinolones versus aerobic and anaerobic isolates from skin and soft tissue samples of human and animal bite wound infections. Antimicrob Agents Chemother 1999; 43: 1475-1479.
59. Goldstein EJC, Citron DM, Hudspeth M, Gerardo SH, Merriam CV. Trovafloxacin compared with levoflaxin, ofloxacin, ciprofloxacinj, azithromycin and clarithromycin against unusual aerobic and anaerobic human and animal bite-wound pathogens. J Antimicrob Chemother 1998; 41: 391-396.
60. Goldstein EJC, Citron DM, Merriam CV, Warren Y, Tyrrel K, Fernandez H. Comparative in vitro activity of ertapenem and 11 other antimicrobial agents against aerobic and anaerobic isolates from skin and soft tissue animal and human bite wound infections. J Antimicrob Chemother 2001; 48: 641-651.
61. Goldstein EJC, Citron DM, Merriam CV, Warren Y, Tyrrel K, Fernandez H. Comparative in vitro activity of faropenem and 11 other antimicrobial agents against 405 aerobic and anaerobic isolates from skin and soft tissue infections from animal and human bites. J Antimicrob Chemother 2002; 50: 411-420.
62. Goldstein EJC, Citron DM, Gerardo SH, Hudspeth M, Merriam CV. Activities of HMR 3004 (RU64004) and HMR 3647 (RU66647) compared to those of Erythromycin, Azythromycin, Clarithromycin, Roxithromycin and eight other antimicrobial agents against unusual aerobic and anaerobic human and animal bite pathogens isolated from skin and soft tissue infections in humans. Antimicrob Agents Chemother 1998; 42: 1127-1132.
63. Goldstein EJC, Citron DM, Merriam CV, Warren Y, Tyrrel K. Comparative in vitro activities of ABT-773 against aerobic and anaerobic pathogens isolated from skin and soft-tissue animal and human bite wound infections. Antimicrob Agents Chemother 2000; 44: 2525-2529.
64. Cercenado E, Cercenado S, Bouza E. In vitro activities of tigecycline (GAR-936) and 12 other antimicrobial agents against 90 Eikenella corrodens clinical isolates. Antimicrob Agents Chemother 2003; 47: 2644-2645.
65. Jaramillo RD, Barraza B, Polo A. Eikenella corrodens. Un modelo biológico de interés para la investigación clínica biomédica y básica. Biociencias 1999; 2: 25-28.