Evaluación inmunohistoquímica de la expresión de calbindina en el cerebro de ratones en diferentes tiempos después de la inoculación con el virus de la rabia
Natalia Lamprea, Biol1, Orlando Torres-Fernández, PhD2
1. Joven Investigadora, Grupo de Microscopía y Análisis de Imágenes, Instituto Nacional de Salud (INS), Bogotá DC, Colombia. e-mail: natalia.lamprea@gmail.com
2. Investigador Científico, Laboratorio de Microscopía, Instituto Nacional de Salud (INS), Bogotá DC, Colombia.
e-mail: otorresf@ins.gov.co
Recibido para publicación diciembre 20, 2007 Aceptado para publicación junio 26, 2008
RESUMEN
Introducción: La calbindina (CB) es una proteína reguladora del metabolismo del calcio intracelular. Previamente se demostró que la infección con virus fijo de la rabia induce pérdida de la expresión de CB, en el cerebro de ratones en estado terminal de la enfermedad, a los 7-8 días de post-inoculación (p.i.) intramuscular.
Objetivo: Determinar si la pérdida de expresión de CB se presenta también en etapas tempranas de la infección y si es una consecuencia inmediata a la aparición de antígenos virales en el cerebro.
Materiales y métodos: Se inocularon ratones con virus fijo de la rabia, por vía intramuscular, en su extremidad posterior izquierda. Diariamente se tomaron entre 4 y 6 animales infectados y sus respectivos controles, se fijaron por perfusión intracardíaca con paraformaldehído, se extrajeron sus cerebros y se obtuvieron cortes coronales en un vibrátomo. Estos se sometieron a reacciones inmunohistoquímicas para evaluar la presencia de antígenos virales y la expresión de CB. Esta última fue cuantificada mediante densitometría óptica en un microscopio con análisis de imágenes.
Resultados: La inmunorreactividad a la rabia fue positiva en el cerebro a partir del día 3 p.i.; se observó primero en neuronas piramidales de la corteza frontal. La inmunotinción para CB no sufrió ninguna alteración cualitativa ni cuantitativa en la corteza cerebral y el estriado hasta el día 6 p.i.
Conclusiones: Estos resultados coinciden con los de otros estudios sobre la rabia en los que se han encontrado alteraciones moleculares, metabólicas o fisiopatológicas en el sistema nervioso sólo en la fase terminal de la enfermedad.
Palabras clave: Rabia; Proteínas de enlace del calcio; Corteza cerebral; Enfermedades cerebrales; Inmunohistoquímica.
Immunohistochemical evaluation of calbindin expression in mouse brain at different times after rabies virus inoculation
SUMMARY
Introduction: The calcium-binding protein calbindin (CB) plays a critical role in intracellular calcium metabolism. In a previous study we demonstrated that the infection with fixed rabies virus caused loss of CB expression in mouse brain during terminal state of the disease, 7-8 days after intramuscular inoculation.
Objectives: To determine loss of CB expression also appears in early stages of the infection and if it is a consequence immediate to the appearance of viral antigens in the brain.
Materials and methods: Mice were inoculated with fixed rabies virus, by intramuscular route, in the left hind limbs. Daily, to sixth day, 4-6 animals and their respective controls were killed and fixed by intracardiac perfusion with paraformaldehyde and their brains were extracted to obtain coronal sections using a vibratome. Free-floating sections were treated by immunohistochemical procedures to evaluate the presence of rabies viral antigens and the expression of CB. The last one was quantified by optical densitometry in a microscope with a system of image analysis.
Results: Rabies immunoreactivity was observed in the brain three days after virus inoculation, beginning in pyramidal neurons of the frontal cortex whereas the immunostaining for CB did not undergo any qualitative nor quantitative changes neither in the cerebral cortex or striatum during the six days post-inoculation.
Conclusions: These results agree with other studies in which it has been settled down that the rabies molecular or metabolic effects on the nervous system are well-known only in the final stage of the disease.
Keywords: Rabies; Calcium-binding proteins; Calbindin; Cerebral cortex; Brain diseases; Immunohistochemistry.
La calbindina (CB) es una proteína muy importante para el mantenimiento de la homeostasis intracelular del calcio1-3. La CB posee un peso molecular aparente de 28 KDa y tiene cuatro sitios de enlace exclusivos a iones Ca+2. Por su alta tasa de unión, así como de afinidad por estos iones, se considera que esta proteína actúa como un tampón de acción rápida2. La CB es dependiente de la vitamina D en tejidos de órganos tales como riñón, intestino y hueso; sin embargo, las concentraciones de CB en el cerebro son independientes del status de la vitamina D3. Esta proteína se encuentra principalmente en el sistema nervioso, en determinadas poblaciones neuronales y su distribución intracelular en el citosol depende de la unión a estructuras citoesqueléticas1-3. En la corteza cerebral la CB se localiza en tres subpoblaciones de interneuronas que corresponden morfológicamente, a células bipenachadas (double bouquet), células neurogliaformes y células de Martinotti4,5. Otras células nerviosas importantes por su contenido de CB son las neuronas espinosas de proyección de tamaño mediano del estriado, muy abundantes en esta estructura, por lo que hacen de ella uno de los referentes neuro-histoquímicos de la presencia de esta proteína6,7. También, parte de las neuronas piramidales del hipocampo y las células granulares del giro dentado exhiben inmunorreactividad notoria a la CB6. Los patrones de distribución de CB son constantes en el tejido cerebral pero se pueden alterar en condiciones patológicas derivadas de enfermedades infecciosas8 o neurodegenerativas9.
Por otra parte, el virus de la rabia infecta casi exclusivamente a las neuronas y una vez invade el sistema nervioso central se desencadenan síntomas clínicos severos y de consecuencias fatales. Sin embargo, los cambios morfológicos macroscópicos y microscópicos que se observan en el tejido cerebral son mínimos10. Por esta razón se ha propuesto que el virus de la rabia da origen a una enfermedad que responde más a cambios bioquímicos en el sistema nervioso que a daño neuronal estructural10,11. Un estudio molecular reveló que la infección experimental en ratones induce la disminución en la expresión de 90% de los genes estudiados mientras que apenas en 1.4% se eleva12. Cualquiera de las dos situaciones puede afectar la regulación del metabolismo celular, la síntesis de proteínas y la integración sináptica.
Las neuronas CB-ir de la corteza cerebral y el cuerpo estriado tienen en común que son células gabaérgicas, es decir, que sintetizan y liberan el neurotransmisor ácido-gamma-aminobutírico (GABA)4,7, por tanto, los cambios en la inmunorreactividad de CB podrían estar relacionados con alteraciones en la neurotransmisión gabaérgica. Existe evidencia experimental del efecto de la rabia sobre el metabolismo del GABA, por esta razón, algunos autores han propuesto que se podría presentar disfunción gabaérgica como parte del cuadro clínico de la rabia13,14. Previamente se ha informado pérdida de CB en el cerebro de ratones, en la fase terminal de la enfermedad, 7-8 días después de inoculación intramuscular con una cepa de virus fijo CVS15,16. El objetivo de este trabajo fue determinar si esta pérdida de CB se inicia en etapas tempranas de la infección y si es consecuencia inmediata a la aparición de antígenos virales en el cerebro.
MATERIALES Y MÉTODOS
Inoculación de ratones. Se utilizaron hembras de ratones ICR (Institute of Cancer Research) de 4 semanas, confinados en la sala de alta seguridad del bioterio del Instituto Nacional de Salud (INS), siguiendo las normas éticas y legales existentes para la investigación con animales de laboratorio. El protocolo de manejo de animales fue aprobado por el Comité de Ética del INS. El inóculo viral [virus fijo de rabia de la cepa CVS (Challenge Virus Standard)] fue suministrado por el Laboratorio de Virología del Instituto Nacional de Salud. Este fue inyectado en un grupo de ratones lactantes, por vía intracerebral, para obtener mayor cantidad de inóculo necesario para el desarrollo del experimento. A partir de un macerado de los cerebros de estos ratones se obtuvo una dilución 1:10 de inóculo viral, en un diluyente que contenía suero equino y antibióticos15 con la que finalmente se realizó la inoculación de los animales objeto de estudio. El virus fue titulado por inoculación intracerebral de diluciones seriadas en ratones (DL50=104). El experimento se llevó a cabo con un total de 25 animales inoculados y sus respectivos controles. La inoculación se realizó por vía intramuscular en la extremidad posterior izquierda, inyectando 0.03 ml de la dilución final (1/10) equivalente a 1000 DL50. Diariamente, a partir del segundo día post-inoculación (p.i.), se tomaron 4-6 animales infectados con sus respectivos controles para sacrificarlos, extraer los cerebros y obtener el material necesario para llevar a cabo el estudio inmunohistoquímico de rabia y CB a medida que progresaba la enfermedad. Los animales se anestesiaron con una solución acuosa de hidrato de cloral (3%, 1 ml/100g de peso) inyectada por vía intraperitoneal, se fijaron por perfusión intracardíaca, inicialmente con solución de tampón de fosfato salino (PBS) y luego con paraformaldehído al 4%. A continuación se extrajeron los cerebros y se post-fijaron en una solución fresca de paraformaldehído al 4% durante 72 horas a 4ºC.
Inmunohistoquímica. Se cortaron rodajas de cerebro de 1 cm de grosor en un plano transversal y se montaron en un vibrátomo para obtener cortes coronales de 100 mm de espesor en dirección rostro-caudal, a nivel del cuerpo calloso y la comisura anterior. Estos cortes se sometieron a las reacciones inmunohistoquímicas, en agitación constante, a temperatura ambiente (20ºC) y con lavados en PBS pH 7.2 después de cada tratamiento. Luego de la fijación, se lavaron los cortes durante toda la noche en PBS y se trataron primero con NH4Cl 0.05 M por 30 min para romper los enlaces aldehído y luego con H2O2 3%, 30 min, para remover la peroxidasa endógena. A continuación se sumergieron los cortes sucesivamente en soluciones de etanol 10%, 20%, 10%, por 5 min en cada una de ellas, para incrementar la permeabilidad del tejido a los anticuerpos. Luego se incubaron, durante una hora, en suero normal de conejo al 3% para bloquear sitios inespecíficos. Se retiró el exceso de suero y a los cortes sin lavar se les agregó la solución que contenía el anticuerpo primario anti-CB (Sigma monoclonal, 1:2500) y se dejaron en la misma durante 20 horas. Después del lavado, los cortes se incubaron durante 2 horas en anticuerpo secundario biotinilado anti-ratón IgG elaborado en conejo (Sigma, 1:600) preparado en PBS con 0.25% de gelatina (Merck) y luego por 2 horas más en la solución del complejo ABC (Vectastain). Para el revelado se utilizó diaminobencidina (DAB) como cromógeno. Para controlar la especificidad de la coloración, y la posible reacción inespecífica del anticuerpo secundario con el tejido, se procesaron cortes de cerebro sin incubación con anticuerpo primario (controles negativos). Un procedimiento similar se llevó a cabo para el estudio inmunohistoquímico de la rabia utilizando un anticuerpo primario elaborado en hamster (dilución 1:160) y anticuerpo secundario anti-hamster (vector, dilución 1:200)17; en este caso el cromógeno utilizado fue el VIP (Vector). Para el control de la técnica inmunohistoquímica en rabia se utilizaron cortes de cerebro no infectado los cuales se procesaron simultáneamente con el material infectado. Para verificar que el virus estuviera presente en la corteza cerebral de cada una de las muestras estudiadas, se obtuvieron cortes seriados que incluían los dos hemisferios, para llevar a cabo, alternativamente, la reacción inmunohistoquímica para la detección de CB o rabia.
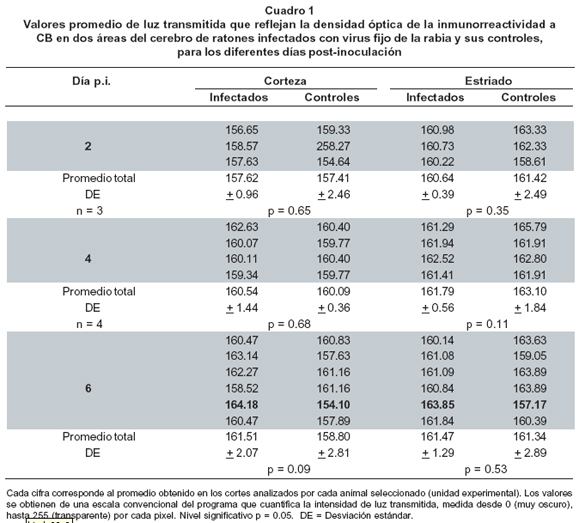
Análisis cuantitativo. Para evaluar cuantitativamente la expresión de la CB en el cerebro de los ratones se seleccionaron las áreas de tejido cerebral con mayor concentración de la proteína: la corteza cerebral frontal supragranular y el cuerpo estriado16. Se cuantificó la densidad óptica del tejido inmunoteñido por medio del programa Zeiss KS-300, en muestras tomadas cada dos días (2, 4 y 6 días p.i.). Se seleccionaron algunos de los cerebros infectados (unidades experimentales), con sus respectivos controles (Cuadro 1). Para cada unidad experimental se seleccionaron 5 cortes y de cada uno de ellos, se tomaron tres campos de lectura de 320 µm x 240 µm para la corteza frontal supragranular y tres para el cuerpo estriado, en áreas vecinas a la comisura anterior. El análisis estadístico se llevó a cabo con una variante de la prueba no paramétrica de Wilcoxon-Mann-Whitney14-16 que utiliza una tabla ideal para experimentos con menos de 10 muestras. La prueba estadística se realizó para cada uno de los tiempos post-infección evaluados.
RESULTADOS
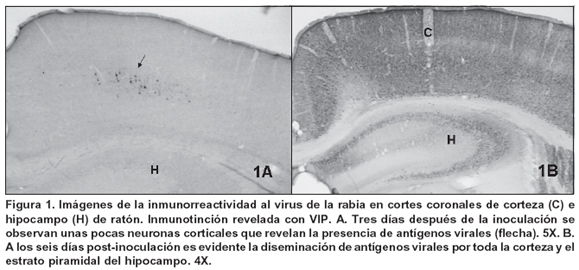
Las primeras neuronas inmunorreactivas a la rabia se observaron a partir del día 3 p.i., en la corteza frontal, en una franja correspondiente a las células piramidales infragranulares, especialmente de la capa V (Figura 1A). Para el día 6, los antígenos virales ya se habían diseminado por todas las áreas corticales y subcorticales y fue notable la inmunorreactividad distribuida por todas las capas neuronales de la corteza cerebral, así como en el estrato de células piramidales del hipocampo (Figura 1B). No obstante, los animales apenas manifestaban signos leves de la enfermedad, tales como movimientos un poco más lentos que los de sus pares control o algún grado de dificultad para mover una de sus extremidades posteriores, por lo general la misma en donde se llevó a cabo la inoculación.
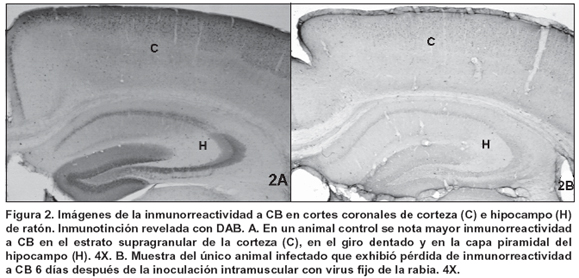
Por otra parte, la inmunorreactividad a CB fue más notoria en la corteza supragranular frontal y parietal, en el cuerpo estriado y en las neuronas piramidales del hipocampo así como en el estrato granular del giro dentado de todas las muestras (Figura 2A). No se observaron diferencias cualitativas ni cuantitativas en las áreas CB-ir al comparar los controles con las muestras de los animales infectados en cada uno de los tiempos p.i. evaluados, con excepción de una muestra de 6 días de infectada en donde la pérdida de inmunotinción de CB fue apreciable (Figura 2B). No obstante, los análisis cuantitativos densitométricos en conjunto para todas las muestras de 6 días de p.i. no arrojaron resultados que demuestren pérdida estadísticamente significativa de CB (Cuadro 1).
DISCUSIÓN
Si bien el virus de la rabia alcanzó rápidamente a la corteza cerebral, la expresión de la proteína CB no parece ser afectada de manera inmediata aún después de que el virus se ha diseminado por casi todas las áreas del cerebro. De acuerdo con estos resultados y estudios previamente publicados15,16, la inmunorreactividad de esta proteína, que refleja su expresión dentro de las neuronas que la portan, se mantiene constante a lo largo del curso de la infección con virus fijo de la rabia y sólo se altera cuando el animal alcanza un estado terminal, es decir, sobre los días 7 y 8 p.i. Un fenómeno similar se informó12 para diferentes genes y proteínas expresados en el SNC del ratón; 90% de los genes analizados no sufrió cambios en la tasa de síntesis o expresión durante el curso de la infección del virus fijo de la rabia pero su expresión disminuyó abruptamente entre los días 6 y 7 p.i. Estos genes están involucrados en el mantenimiento de las funciones normales de las células.
Schneider escribió18: «Los cambios patológicos en la rabia aparecen tarde durante la infección y no se pueden correlacionar con el contenido viral de los tejidos». El estudio de otros aspectos de la infección por rabia coincide en demostrar alteraciones tardías. Así, por ejemplo, al evaluar la actividad eléctrica cerebral, en ratones infectados con virus �calle�, los patrones del electroencefalograma (EEG) se preservan sin cambios, durante casi toda la duración de la enfermedad, hasta que el EEG colapsa pocas horas antes de la muerte. Igualmente tardía es la respuesta inmunológica. Los títulos de anticuerpos virales específicos son por lo general bajos en las infecciones humanas hasta sólo unas horas antes de la muerte19. Todo esto se resume en que el virus permite la preservación de la integridad de la red neuronal hasta bien avanzada la infección como estrategia de supervivencia y para facilitar su transmisión20.
Este comportamiento de la infección por VR hace pensar en dos cosas importantes:
AGRADECIMIENTOS
A COLCIENCIAS y a la Subdirección de Investigación del Instituto Nacional de Salud por la financiación del proyecto código 2104-04-18104. Al Dr. Jaime Castellanos, Director del Instituto de Virología de la Universidad El Bosque, por su colaboración para la titulación del virus de la rabia.
REFERENCIAS
1. Schwaller B, Meyer M, Schiffmann S. «New» functions for «old» proteins: The role of the calcium-binding proteins calbindin D-28k, calretinin and parvalbumin, in cerebellar physiology. Studies with knockout mice. Cerebellum. 2002; 1: 241-58.
2. Lutz W, Frank EM, Craig T, Thompson R, Venters RA, Kojetin D, et al. Calbindin D28K interacts with ran-binding protein M: identification of interacting domains by NMR spectroscopy. Biochem Biophys Res Commun. 2003; 303: 1186-92.
3. Zheng W, Xie Y, Li G, Kong J, Fena JQ, Li YC. Critical role of calbindin-D28k in calcium homeostasis revealed by mice lacking both vitamin D receptor and calbindin-D28k. J Biol Chem. 2004; 279: 52406-13.
4. DeFelipe J. Types of neurons, synaptic connections and chemical characteristics of cells immunoreactive for calbindin-D28K, parvalbumin and calretin in the neocortex. J Chem Neuroanat. 1997; 14: 1-19.
5. Sun XZ, Takahashi S, Cui C, Inoue M, Fukui Y. Distribution of calbindin immunoreactive neurons in rat primary motor cortex. J Med Invest. 2002; 49: 35-9.
6. Celio MR. Calbindin D-28k and parvalbumin in the rat nervous system. Neuroscience. 1990; 35: 375-475.
7. Pickel VM, Heras A. Ultrastructural localization of calbindin-D28K and GABA in the matrix compartment of the rat caudate-putamen nuclei. Neuroscience. 1996; 71: 167-78.
8. Masliah E, Ge N, Achim CL, Wiley CA. Differential vulnerability of calbindin-immunoreactive neurons in HIV encephalitis. J Neuropathol Exp Neurol. 1995; 54:350-7.
9. Geula C, Bu J, Nagykery N, Scinto L, Chan J, Joseph J, et al. Loss of calbindin from aging human cholinergic basal forebrain: Relation to neural loss. J Comp Neurol. 2003; 455: 249-59.
10. Iwasaki Y, Tobita M. Pathology. In: Jackson AC, Wunner WH, editores. Rabies. San Diego: Academic Press; 2002. p. 283-306.
11. Fu ZF, Jackson AC. Neuronal dysfunction and death in rabies virus infection. J Neurovirol. 2005; 11: 101-16.
12. Prosniak M, Hooper C, Dietzschold B, Koprowsky H. Effect of rabies virus infection on gene expresión in mouse brain. Proc Natl Acad Sci. USA 2001; 98: 2758-63.
13. Ladogana A, Bouzamondo E, Pocchiari M, Tsiang H. Modification of tritiated gama amino-butyric acid transport in rabies virus-infected primary cortical cultures. J Gen Virol. 1994; 75: 623-7.
14. Rengifo AC, Torres-Fernández O. Disminución del número de neuronas que expresan GABA en la corteza cerebral de ratones infectados con rabia. Biomedica. 2007; 27: 548-58.
15. Torres-Fernández O, Yepes GE, Gómez JE, Pimienta HJ. Efecto de la infección por el virus de la rabia sobre la expresión de parvoalbúmina, calbindina y calretinina en la corteza cerebral de ratones. Biomedica 2004; 24: 63-78.
16.Torres-Fernández O, Yepes GE, Gómez JE, Pimienta HJ. Calbindin distribution in cortical and subcortical brain structures of normal and rabies-infected mice. Int J Neurosci. 2005; 115: 1375-82.
17.Sarmiento L, Rodríguez G, de Serna C, Boshell J, Orozco L. Detection of rabies virus antigens in tissue: immunoenzimatic method. Patologia. 1999; 37: 7-10.
18.Schneider L. Spread of virus within the central nervous system. In: Baer G, editor. The natural history of rabies. Vol. 1. New York: Academic Press; 1975. p. 199-216.
19.Tsiang H. Pathophysiolgy of rabies virus infection of the nervous system. Adv Virus Res. 1993; 42: 375-412.
20. Lafon M. Modulation of the immune response in the nervous system by rabies virus. Curr Top Microbiol Immunol. 2005; 289: 239-58.
LISTA ABREVIATURAS
Nota: Los productos ABC y VIP son marcas registradas y no corresponden a siglas