Evaluación de la expresión del factor de transcripción asociado con microftalmia (MITF) en sangre periférica de individuos con y sin melanoma maligno y en líneas celulares*
Nelson Rangel, MSc1, Milena Rondón, MSc1, Sandra Ramírez, PhD1
* Este trabajo fue realizado con el soporte del Fondo de Investigaciones de la Universidad del Rosario (FIUR), aprobado en el año 2003.
1. Laboratorio de Biología Celular y Molecular, Instituto de Ciencias Básicas, Facultad de Medicina, Universidad del Rosario, Bogotá DC, Colombia. e-mail: nelsonrangel06@urosario.edu.co sandra.rondonla@urosario.edu.co sramire@urosario.edu.co
Recibido para publicación diciembre 7, 2007 Aceptado para publicación enero 15, 2009
RESUMEN
Introducción: La incidencia de melanoma maligno se ha incrementado más rápido que cualquier otro tipo de cáncer, intensificando así la búsqueda de herramientas que faciliten la identificación temprana del melanoma. El factor de transcripción asociado con microftalmia (MITF) es conocido como el regulador maestro de melanocitos. En el presente estudio se analiza la expresión del gen MITF en sangre periférica de un grupo de individuos con melanoma, comparándola con un grupo de personas sin cáncer y en algunas líneas celulares.
Materiales y métodos: Se extrajo ARN de 31 muestras de sangre periférica: 19 de pacientes con melanoma y 12 de personas sin ningún tipo de cáncer. Se cuantificaron niveles de expresión tanto para el gen MITF como para los genes de expresión constitutiva (b2M y GAPDH) mediante PCR tiempo real. Asimismo se evaluó la expresión de los mismos genes en cinco líneas celulares.
Resultados: En todos los individuos se observó expresión del gen MITF, aunque no hubo diferencias estadísticamente significativas entre los niveles de expresión en los grupos de estudio (p=0.09). Sin embargo, la expresión de MITF en el grupo de pacientes con melanoma fue más variable que la observada en el grupo de personas sin cáncer. Asimismo, en la línea celular de adenocarcinoma gástrico se detectó expresión del gen MITF, no descrita hasta el momento.
Conclusiones: Se encontraron niveles de expresión del gen MITF en sangre periférica tanto de personas con melanoma como en personas sin cáncer. Sin embargo, la variabilidad en los niveles de expresión del gen MITF observados en personas con melanoma, sugiere la posible presencia de células tumorales en circulación.
Palabras clave: Melanoma; MITF; qPCR; Células tumorales en circulación; Cáncer.
Evaluation of microphthalmia associated transcription factor (MITF) expression in peripheral blood of a population with malign melanoma and control population and cell lines
SUMMARY
Background: The incidence of malign melanoma tumours has increased more rapidly than any other type of cancer; this has intensified the search for tools that facilitate early identification of melanoma. Microphthalmia associated transcription factor (MITF) is currently known as being a master melanocyte regulator; we analyse MITF gene expression in peripheral blood from individuals suffering from melanoma, compared to people without any type of cancer and ones cell lines.
Materials and methods: Thirty one samples of peripheral blood were used: 19 from patients having melanoma and 12 from people without any cancer. RNA was then extracted from these samples. MITF and housekeeping genes (b2M and GAPDH) expression levels were then quantified by real-time PCR. Five cell lines were also used to determine the MITF expression
Results: MITF gene expression could be observed in all individuals, though no statistically significant differences were found among expression levels in the groups studied (p=0.09). Even so, MITF expression in the group of patients suffering from melanoma was much more variable than that observed in the group of cancer-free people. Expression was detected in the cell line AGS (gastric adenocarcinoma), not yet described.
Conclusions: MITF gene expression levels were detected in the peripheral blood of both people suffering from melanoma and people without any type of cancer. However, variability in the number of molecules in MITF gene expression was observe in people with melanoma, this suggest the presence of tumour cells in circulation.
Keywords: Melanoma; MITF; qPCR;Circulation tumor cells; Cancer.
El melanoma maligno es un tipo de cáncer considerado en algunos países (Estados Unidos y Australia) como un problema de salud pública, al extremo de catalogarse epidemia, debido a que en los últimos años la incidencia de la enfermedad a aumentado1. Está considerado entre los tipos de cáncer más agresivos y en Colombia es la causa de aproximadamente 40% de las muertes por cáncer de piel2, por lo cual se hace importante implementar pruebas de detección temprana, pronóstico y seguimiento basadas en marcadores moleculares, además de orientar y facilitar el desarrollo de nuevas opciones de tratamiento. Actualmente la atención se centra en la detección de células tumorales circulantes en sangre periférica, como herramienta promisoria para la identificación ya sea de personas con estadíos tempranos de la enfermedad y/o con resultados clínicos potencialmente pobres.
Smith et al.3 fueron los primeros en sugerir que células de melanoma podrían ser detectadas en sangre periférica al identificar moléculas de ARNm específicas de células tumorales circulantes (ARNm de tirosinasa), pero ausentes en células normales no malignas. A partir de este informe original varios grupos de investigación han utilizado ésta y otras metodologías para detectar la expresión de genes específicos de melanocitos en circulación (Tirosinasa, Dct, Mart-1)4,5. Sin embargo, en otros estudios se argumenta que estos genes no son candidatos suficientemente buenos para el análisis, sobre todo en estados avanzados de la enfermedad, porque sus moléculas de ARNm no son detectadas de manera constante en sangre periférica de individuos con melanoma6.
En cuanto al factor de transcripción asociado con microftalmia (MITF) se ha demostrado que cumple un papel fundamental en la diferenciación y proliferación, así como en la viabilidad post-natal (sobrevida) de los melanocitos porque regula la expresión de genes como p21, INK4a, CDK2, BCL2, TYR, DCT, entre otros, al ser activado principalmente a través de las vías de señalización MAPK, cAMP/PKA/CREB y WNT-b catenina7,8. Han sido descritos en el gen MITF 9 promotores diferentes, cada uno de los cuales codifica una isoforma con extremo 5' único, aunque tienen en común los exones 2 al 9 que codifican para los dominios funcionales de los factores de transcripción. Se ha informado la presencia de algunas de las isoformas de manera tejido-específica como es el caso de la isoforma MC, expresada específicamente en mastocitos y la isoforma M expresada en melanocitos9. Además, técnicas de inmunohistoquimica en tejido tumoral han mostrado que la expresión del gen MITF, a diferencia de otros marcadores específicos de células pigmentadas, se mantiene en muchos, sino en todos los tumores de origen melanocítico, estableciéndolo como un marcador lo suficientemente sensible para tenerlo en cuenta como posible herramienta que complemente las que en la actualidad se usan para el diagnóstico del melanoma y relevando su importancia en el desarrollo de la enfermedad10,11. Por tal motivo en el presente estudio analítico de casos y controles, se evaluó la expresión del gen MITF en sangre periférica de individuos con melanoma maligno (estadíos III y IV), y se comparó con la expresión en sangre periférica de personas sin ningún tipo de cáncer mediante PCR tiempo real (qPCR) y en diferentes líneas celulares.
MATERIALES Y MÉTODOS
Cultivos celulares. Cinco líneas celulares se utilizaron para evaluar la expresión del gen MITF. La línea celular A-375 (melanoma maligno) utilizada como control positivo, fue obtenida directamente de la ATCC (American Type Culture Collection) mientras que las líneas celulares AGS y MCF-7 (adenocarcinoma gástrico y cáncer de mama, respectivamente) fueron donadas por la unidad de Inmunología del Instituto Nacional de Cancerología (INC), Bogotá, Colombia. Las líneas celulares se mantuvieron en cultivo según las especificaciones del proveedor. En general, el medio de cultivo DMEM se suplementó con suero fetal bovino (SFB) al 10% y se incubaron a 37ºC con una atmósfera de 5% de CO2. Se generaron cultivos primarios de melanocitos (CPM) y fibroblastos (CPF) a partir de biopsias de piel, los cuales se sometieron a la acción de la solución Dispasa II (Roche Molecular Biochemicals) al 0.5%. Los fragmentos disgregados de epidermis (melanocitos) se colocaron en caja de Petri estéril con 2 ml de medio de crecimiento específico de melanocitos (Cell Application) y los fragmentos de dermis (fibroblastos) en caja de Petri estéril con 2 ml de medio DMEM enriquecidos con 10% de SFB. Las células obtenidas se incubaron a 37ºC y 5% de CO2, hasta que alcanzaron una confluencia de 70%, momento en el cual se trataron con tripsina EDTA (0.25% por 60 seg) para desprender las células y usando el reactivo TRIZOL para preparar los extractos celulares con el fin de aislar el ARN total.
Selección de casos y controles. Teniendo en cuenta la incidencia de la enfermedad en la población del presente estudio e informes previos5, se calculó el tamaño de muestra representativo a ser tenido en cuenta, utilizando el programa tamaños de muestra 1.1. Se reclutaron para participar en el estudio 13 hombres y 6 mujeres entre 13 y 86 años de edad; 19 con melanoma maligno diagnosticados clínica e histológicamente de acuerdo con el AJCC (American Joint Comité on Cancer), 12 con metástasis a nódulos linfáticos regionales (estadío III) y 7 con metástasis distantes (estadío IV). Todos fueron diagnosticados, tratados y seguidos en las Unidades de Dermatología y Seno y Tejidos Blandos del Instituto Nacional de Cancerología (INC), Bogotá, Colombia. Se excluyeron del estudio los pacientes que recibieron terapia sistémica anti-neoplásica (quimio o radioterapia). Se incluyeron como controles 12 personas (8 hombres y 4 mujeres) sin ningún tipo de cáncer que cumplieron con características de género, edad, ocupación y condición socioeconómica semejantes a las de los pacientes, con el fin de realizar análisis pareados y controlar posibles variables de confusión como lo es la exposición a radiación ultravioleta. Todos los individuos participantes firmaron consentimiento informado aprobado por los comités de ética de la Universidad del Rosario y del INC, de acuerdo con lo establecido en la declaración de Helsinki.
Recolección y procesamiento de muestras sanguíneas. Se recolectaron en tubos vacutainer con EDTA muestras de sangre (5 ml) de pacientes con melanoma y personas sin ningún tipo de cáncer y se procesaron en máximo 2-3 h después de la recolección. Se separaron los glóbulos blancos y otras células mononucleares utilizando un gradiente de Ficoll Histopaque-1077 (Sima Aldrich) y centrifugando a 2000 rpm por 20 min. Se aisló ARN total de células mononucleares utilizando TRIzol (InvitrogenTM, Life Technologies), técnica que se fundamenta en el método de extracción por fenol-cloroformo-isotiocianato de guanidina. La concentración, pureza e integridad del ARN total obtenido fue determinada por espectrofotometría y visualizado por electroforesis en gel de agarosa denaturante al 1% para la posterior síntesis del ADNc.
RT-PCR (síntesis de ADN complementario-ADNc). Se sometió ARN total a digestión enzimática con la ADNsa I grado amplificación (InvitrogenTM, Life Technologies), para eliminar trazas de ADN genómico. Para cada una de las muestras incluidas en el estudio, dos reacciones de RT-PCR (Reverse Transcription Polymerase Chain Reaction), se llevaron a cabo con el Kit SuperScriptTMFirst-Strand Synthesis System (InvitrogenTM, Life Technologies). Se sintetizó ADNc siguiendo las instrucciones del proveedor. Brevemente, se mezcló 1 µg de ARN total con 0.4 mM de dNTPs, 20 ng oligo (dT) y agua DEPC.
La solución se sometió a un paso de denaturación, calentando a 65ºC por 5 min. Luego se completó a un volumen final de 25µl agregando buffer RT 1X, 4 mM MgCl2, 8 mM DTT, 40 U RnaseOUTTM y 50 U SuperScriptTMII. La mezcla de reacción se incubó a 42ºC por 50 min y se bloqueó la actividad polimerasa aumentando la temperatura a 70ºC por 15 min. 2U de RNasa H se adicionaron a la mezcla, la cual se incubó a 37º por 20 min para eliminar restos de ARN contaminante.
PCR tiempo real (qPCR). Mediante PCR tiempo real, con cebadores específicos de cada gen se amplificaron regiones del ADNc para evaluar la expresión de los genes GAPDH, b2M y MITF utilizando el tinte fluorescente de unión a ADN de doble hebra SYBR Green ITM en el detector de fluorescencia Opticon II (MJ Research). Se llevaron 4 µl del ADNc obtenido en la RT-PCR, a un volumen final de 20 µl usando la mezcla de reacción del Kit Platinum® SYBR® Green qPCR SuperMix UDG 2X (InvitrogenTM, Life Technologies). La concentración final de los componentes en la mezcla para la amplificación por PCR tiempo real fue: Master Mix 1X, (1.5U Platinum® Taq DNA Polimerasa, 20 mM Tris-HCl (pH 8.4), 50 mM KCl, 3 mM MgCl2, 200 mM c/u dNTPs, 1U UDG) y 0.5 µM de c/u de los primers (Cuadro 1)12 en H2O destilada estéril. Las condiciones de PCR utilizadas fueron las mismas para los tres genes: 50°C por 3 min, 94°C por 15 min, 40 ciclos a 95°C por 15 seg, 58°C por 30 seg, Plate Read (Lectura de Fluorescencia) y 72°C por 30 seg, luego 72°C por 5 min y por último se obtuvo la curva de disociación entre 70ºC y 90°C. Los productos amplificados de los genes b2M y GAPDH, se purificaron con el Kit QIAquick® PCR Purification (Quiagen), cuantificados por espectrofotometría (TECAN® Genios) y utilizados directamente para realizar diluciones y generar curvas estándar en el equipo de PCR tiempo real. El producto de PCR del gen MITF fue purificado y clonado en el vector plasmídico TOPO 2.1 (InvitrogenTM, Life Technologies) para generar un producto más estable y específico a utilizar en las curvas estándar.
Análisis estadístico. Los niveles de expresión entre los grupos estudiados se compararon utilizando análisis de cuantificación relativa con el método publicado por Pfaffl, M13. Asimismo, análisis de cuantificación absoluta para determinar el número de moléculas permitieron establecer normalidad en la distribución de los datos, aplicando la prueba Shapiro-Wilks. Diferencias significativas entre los grupos estudiados para los niveles de expresión en cuanto al número de moléculas de los genes MITF, GAPDH y b2M fueron evaluadas utilizando las pruebas no paramétricas Kruskal-Wallis y la prueba de rangos con signo de Wilcoxon. Todas las pruebas se evaluaron con un nivel de significancia del 95% (p=0.05) en el programa estadístico SPSS (Versión 12).
RESULTADOS
La población de casos que inicialmente se incluyó en el estudio fue de 19 personas con melanoma maligno, cuya característica principal era el estadío clínico de la enfermedad (III y IV). En los ensayos de PCR tiempo real realizados para los genes GAPDH y b2M se detectó amplificación de los mismos en todas las muestras estudiadas. Sin embargo, 7 casos se excluyeron porque en el análisis de exploración con gráficos de caja y bigote (datos no mostrados), se observaron valores atípicos y atípicos extremos indicando que la integridad y calidad de estas muestras fue baja y además podrían generar resultados erróneos en el análisis final. Como se hizo un equiparamiento 1:1 por sexo, edad, estrato y ocupación, los análisis estadísticos pareados se realizaron con 12 pacientes con melanoma maligno y 12 personas sin ningún tipo de cáncer. La cantidad de ARN obtenido a partir de este grupo de muestras varió entre 10 y 15 µg por muestra, usando por ensayo 1 µg. Asimismo, también se calculó la absorbancia a 280 nm y su relación con la absorbancia 260 nm (260/280) proporcionó datos superiores a 1.7, indicando la buena calidad y pureza del material extraído.
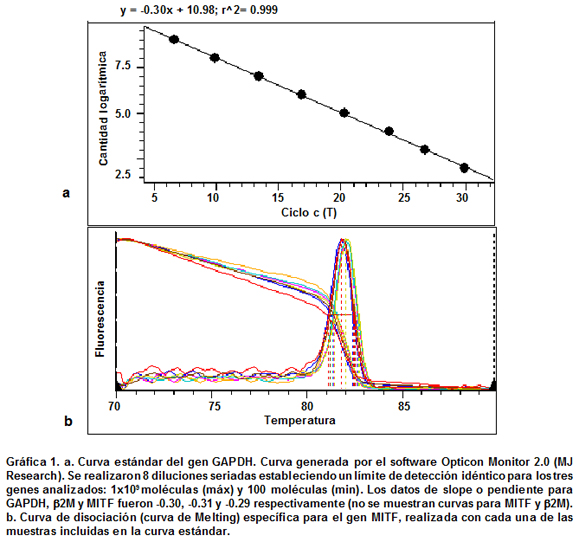
Curvas estándar y análisis de eficiencia. Los métodos de cuantificación (absoluta y relativa), requieren que las eficiencias de PCR de todos los genes sean iguales o muy similares con un nivel bajo de variación (preferiblemente entre 90% y 100% de eficiencia), con el fin de obtener datos reales y confiables para los análisis de cuantificación14. Los productos purificados y cuantificados de los genes GAPDH y b2M, así como el producto clonado del gen MITF, se utilizaron para generar ocho diluciones seriadas (1/10) incluidas en cada una de las curvas estándar. El límite de detección para todas las curvas se ubicó entre 1x109 y 100 moléculas que corresponden al menor y mayor C(t) (Cycle threshold) detectados, respectivamente (Gráfica 1a). La eficiencia en la amplificación de los tres genes estudiados se ubicó entre 95% y 100% y se analizó utilizando la fórmula E = [10(-1/-S)] donde S representa el valor de la pendiente o slope de la curva. Además, los análisis de curva de melting en el rango de temperatura entre 70ºC y 90°C permitieron determinar la especificidad de los primers, porque sólo se observó un pico o punto de melting para cada uno de los genes (Gráfica 1b).
Cuantificación del gen MITF en líneas celulares. En las líneas celulares (A-375, AGS, MCF-7) y en los cultivos primarios (CPM, CPF) se obtuvo producto de amplificación cuando se usaron cebadores específicos de los genes GAPDH y b2M. Por lo menos se realizaron dos experimentos independientes por duplicado para cada cultivo, obteniendo datos muy similares, que aseguraron la reproducibilidad de los ensayos de PCR tiempo real. Los datos entre cultivos y entre genes, mostraron semejanzas importantes en sus C(t)s, lo cual permitió establecer inicialmente a GAPDH y b2M como genes que se expresan de forma relativamente constante en los cultivos celulares analizados (datos no mostrados).
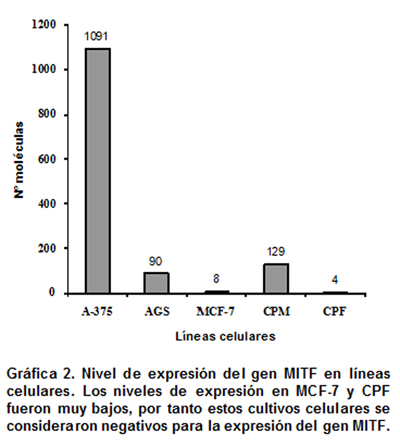
En la línea celular de melanoma maligno A-375 y en el cultivo primario de melanocitos (CPM), se pudo detectar la expresión del gen MITF. En la línea celular de cáncer gástrico (AGS), también se evidenció expresión del gen MITF, hecho inesperado, ya que estas células son de origen epitelial gástricas en las que no se esperaba encontrar expresión del mismo y para las cuales no existe ningún tipo de informe con respecto a la expresión del gen evaluado. Los niveles de expresión detectados en el cultivo primario de fibroblastos (CPF) y en la línea celular de cáncer seno (MCF-7) fueron bastante bajos, ubicándose fuera del límite de detección, por lo que se consideraron negativos para la expresión del gen MITF (Gráfica 2).
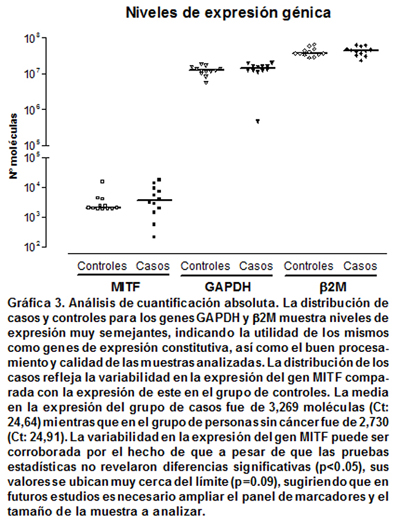
Cuantificación del gen MITF en casos y controles. Al realizar análisis de cuantificación absoluta, la media calculada para los niveles de la expresión del gen GAPDH se ubicó entre 10 y 12 millones de moléculas, tanto para el grupo de casos como para el grupo de controles. Por otro lado la media en la expresión del gen b2M se ubicó entre 41 y 43 millones de moléculas tanto para los pacientes con melanoma maligno como para las personas sin cáncer (Gráfica 3). Como los controles fueron seleccionados teniendo en cuenta características de género, edad, ocupación y condición socioeconómica semejantes a las de los pacientes, los análisis se realizaron de forma pareada aplicando la prueba no paramétrica de rangos con signo de Wilcoxon, con la cual no se encontraron diferencias significativas en los niveles de expresión de los genes GAPDH (p=0.388) y b2M (p=0.347) entre las parejas de los grupos de estudio.
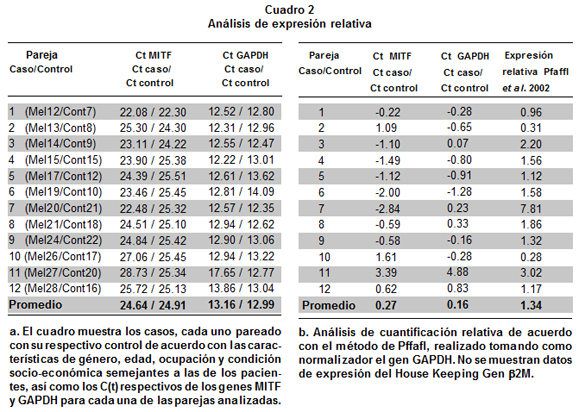
Análisis de cuantificación absoluta permitieron detectar niveles de expresión del gen MITF en todos los individuos con melanoma y en todos los individuos sin cáncer, sin embargo se observó mayor variabilidad en los niveles de expresión en el grupo de casos, hecho que es corroborado al comparar la dispersión de sus muestras con respecto a la dispersión de las muestras del grupo control (Gráfica 3). Los niveles de expresión observados para cada uno de los grupos evaluados, se ubicaron entre 2,700 y 3,300 moléculas. No se encontraron diferencias significativas (p>0.05) en los niveles de expresión del gen MITF entre pacientes con melanoma y personas sin ningún tipo de cáncer, al aplicar las pruebas estadísticas con los datos crudos de C(t) (Wilcoxon p= 0.38). De igual forma, no se encontraron diferencias significativas al comparar el número de moléculas del gen MITF, entre casos y controles (p>0.05). Sin embargo, se reflejan pequeñas diferencias en el hecho de que en 8 de las 12 parejas analizadas los casos mostraron mayores niveles de expresión (Cuadro 2a, Gráfica 3). Análisis de cuantificación relativa realizados con el método publicado por Pfaffl13, el cual tiene en cuenta la eficiencia de amplificación tanto del gen blanco (MITF) como la del gen normalizador (GAPDH), permitieron evidenciar que en promedio, la diferencia en el nivel de expresión del gen MITF fue de 1.34 veces más en el grupo de casos respecto al grupo control (Cuadro 2a y 2b), confirmando las pequeñas diferencias encontradas en el análisis de cuantificación absoluta.
DISCUSIÓN
La detección de células tumorales en sangre periférica de pacientes con cáncer es un procedimiento que ha sido ampliamente utilizado para identificar moléculas de ARNm específicas de tumor en varios tipos de malignidad como por ejemplo en el cáncer de próstata, cáncer de seno y melanoma maligno15. Sin embargo, se presentan dificultades para detectar y cuantificar dicho ARNm, particularmente por la gran variabilidad y falta de sensibilidad de las técnicas empleadas, siendo la RT-PCR una de las técnicas con mayor grado de sensibilidad y especificidad, motivo por el cual se utiliza con frecuencia para análisis de expresión génica. Asimismo, el biomarcador más utilizado para detectar células tumorales en circulación de pacientes con melanoma, la tirosinasa (enzima clave para la biosíntesis de melanina), presenta variaciones considerables en la detección del mismo que van desde 0%16 hasta 100%17, lo cual hace necesaria la búsqueda de otros marcadores en los que se observe menor variabilidad. A diferencia de los estudios citados en el presente trabajo se usó la PCR cuantitativa (qPCR) para detectar los niveles de expresión del gen MITF en sangre periférica de pacientes con melanoma maligno como indicador de la presencia de células tumorales en circulación.
El análisis cuantitativo de la expresión del gen MITF se realizó aplicando los métodos de cuantificación absoluta y relativa, basados en la generación de curvas estándar. Estas se crearon a partir de un vector plasmídico que contenía el inserto de 124pb correspondientes a la secuencia consenso del gen (melanocitos expresan otras isoformas además de MITF-m).
Análisis en cultivos celulares, revelaron niveles de expresión del gen MITF en la línea celular de melanoma maligno (A-375) y en el cultivo primario de melanocitos (CPM). En la línea celular de cáncer de seno (MCF)-7 y en el cultivo primario de fibroblastos (CPF), se detectaron señales de amplificación muy tardías, datos que fueron práctica- mente no cuantificables y por tanto considerados negativos para la expre-sión del gen MITF. La reproducibilidad de los datos, obtenida de 2 experimentos independientes por triplicado de estos cultivos celulares (MCF-7 y CPF), podría explicarse por transcripción ilegítima del gen, reportada en algunas investigaciones18 como la responsable de generar niveles muy bajos de expresión de cualquier gen en cualquier tipo celular. En la línea celular de cáncer gástrico (AGS) un promedio de 90 moléculas fue detectado después de realizar 2 experimentos independientes por triplicado. Poco o nada ha sido informado sobre la expresión de MITF en el tejido gástrico, por tanto estos datos se constituyen en el primer reporte de la expresión del gen en esta línea celular y posiblemente el primero en sugerir la expresión de una o más isoformas del gen MITF en el tejido gástrico.
Se encontró expresión del gen MITF tanto en los 12 pacientes con melanoma maligno (estadíos III-IV) como en las 12 personas sin ningún tipo de cáncer. El análisis de cuantificación relativa mostró que en promedio los casos expresaron 1.34 veces más el gen MITF con respecto al grupo de personas sin cáncer; en el análisis de cuantificación absoluta al comparar los datos obtenidos (número de moléculas) de manera pareada (caso-control), no se encontraron diferencias estadísticamente significativas en los niveles de expresión del gen MITF, entre los grupos evaluados (p>0.05). Se descartaron estimaciones erróneas durante el proceso de cuantificación porque tampoco se encontraron diferencias significativas en los genes de expresión constitutiva GAPDH y b2M (p>0.05), indicando que los procedimientos se realizaron correctamente. MITF es esencial para el desarrollo y sobrevida de melanocitos y al parecer también cumple este importante papel en otros tipos celulares al codificar isoformas que tienen diferentes extremos amino-terminales. Una de ellas es MITF-A expresada en mastocitos19, células reconocidas por ser efectores principales de reacciones alérgicas e inflamatorias, derivadas de células madre hematopoyéticas. La circulación normal de estas células en el torrente sanguíneo, así como la identificación de la proteína MITF en análisis inmunohistoquímicos en linfocitos20, podrían explicar la detección del gen tanto en personas con melanoma maligno como en personas sin ninguna tipo de cáncer y de la misma forma justificar por qué no se encontraron diferencias significativas entre los grupos estudiados. A pesar de que no se observaron diferencias significativas en la cuantificación absoluta, pequeñas diferencias (p=0.09) son reflejadas por la amplia variabilidad en los niveles de expresión del gen MITF (Gráfica 3), así como también por el aumento pequeño en el nivel de expresión de los casos observado al realizar análisis de cuantificación relativa (Cuadro 2b), sugiriendo que puede existir una mayor inestabilidad genética en las células circulantes del grupo de casos, hecho confirmado por estudios de citogenética en nuestro grupo (artículo en preparación), la cual podría explicarse por la presencia de células tumorales en sangre periférica de pacientes con melanoma.
Las similaridades en los niveles de expresión del gen MITF entre los grupos estudiados, sugiere que este gen es ampliamente expresado, indicando que puede ser un gen de expresión constitutiva que poco puede contribuir en el diagnóstico específico del melanoma, tal como lo informan algunos autores10,11. Sin embargo, se debe tener en cuenta que mientras la expresión del gen MITF en personas sin cáncer fue relativamente constante, en el grupo de personas con melanoma la expresión observada fue mucho más variable. Por tanto, es posible que ampliando el número de individuos a analizar y utilizando un panel de biomarcadores que incluyan por ejemplo la tirosinasa y la isoforma específica de melanocitos del gen MIFT (MITF-m), se pueda obtener una mayor especificidad y sensibilidad para desarrollar y validar pruebas de diagnóstico molecular, que permitan detectar células tumorales en circulación de pacientes con melanoma.
AGRADECIMIENTOS
Los autores agradecen a los Servicios de Dermatología y Seno y Tejidos Blandos del Instituto Nacional de Cancerología (INC), en especial a los doctores Álvaro Acosta, Javier Rueda y Fernando Perry así como al doctor William Sánchez del Hospital Militar Central por su colaboración en el contacto y recolección de los pacientes. También agradecen a los doctores Pablo Moreno y Ricardo Cendales del INC por su gran ayuda en el desarrollo del diseño experimental y análisis estadísticos, respectivamente.
REFERENCIAS
1. Barzilai DA, Singer ME. The potential impact on melanoma mortality of reducing rates of suboptimal excision margins. J Invest Dermatol. 2003; 120: 1067-72.
2. Pardo C. Casos nuevos de cancer en el Instituto Nacional de Cancerología, Colombia. Rev Colomb Cancerol 2002; 7: 4-19.
3. Smith B, Selby P, Southgate J, Pittman K, Bradley C, Blair GE. Detection of melanoma cells in peripheral blood by means of reverse transcriptase and polymerase chain reaction. Lancet. 1991; 338: 1227-9.
4. Keilholz U, Goldin-Lang P, Bechrakis NE, Max N, Letsch A, Schmittel A, et al. Quantitative detection of circulating tumor cells in cutaneous and ocular melanoma and quality assessment by real-time reverse transcriptase-polymerase chain reaction. Clin Cancer Res. 2004; 10: 1605-12.
5. Samija M, Juretic A, Solaric M, Samija I, Bingulac-Popovic J, Grahovac B, et al. RT-PCR detection of tyrosinase, gp100, MART1/Melan-A, and TRP-2 gene transcripts in peripheral blood of melanoma patients. Croat Med J. 2001; 42: 478-83.
6. Farthmann B, Eberle J, Krasagakis K, Gstottner M, Wang N, Bisson S, et al. RT-PCR for tyrosinase-mRNA-positive cells in peripheral blood: evaluation strategy and correlation with known prognostic markers in 123 melanoma patients. J Invest Dermatol. 1998; 110: 263-7.
7. Levi CK, Fisher MD. MITF: master regulator of melanocyte development and melanoma oncogene. Trends Mol Med. 2006; 12: 14.
8. Vance KW, Goding CR. The transcription network regulating melanocyte development and melanoma. Pigment Cell Res. 2004; 17: 318-25.
9. Hershey CL, Fisher DE. Genomic analysis of the microphthalmia locus and identification of the MITF-J/Mitf-J isoform. Gene. 2005; 347: 73-82.
10. Carr KM, Bittner M, Trent JM. Gene-expression profiling in human cutaneous melanoma. Oncogene. 2003; 22: 3076-80.
11. Miettinen M, Fernández M, Franssila K, Gatalica Z, Lasota J, Sarlomo-Rikala M. Microphthalmia transcription factor in the immunohistochemical diagnosis of metastatic melanoma: comparison with four other melanoma markers. Am J Surg Pathol. 2001; 25: 205-11.
12. Bustin SA. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J Mol Endocrinol. 2000; 25: 169-93.
13. Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001; 1: 29-45.
14. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2 [-Delta Delta C(T)] method. Methods. 2001; 25: 402-8.
15. Ghossein RA, Bhattacharya S, Rosai J. Molecular detection of micrometastases and circulating tumor cells in solid tumors. Clin Cancer Res. 1999; 5: 1950-60.
16. Foss AJ, Guille MJ, Occleston NL, Hykin PG, Hungerford JL, Lightman S. The detection of melanoma cells in peripheral blood by reverse transcription-polymerase chain reaction. Br J Cancer. 1995; 72: 155-9.
17. Brossart P, Keilholz U, Willhauck M, Scheibenbogen C, Mohler T, Hunstein W. Hematogenous spread of malignant melanoma cells in different stages of disease. J Invest Dermatol. 1993; 101: 887-9.
18. Chelly J, Concordet JP, Kaplan JC, Kahn A. Illegitimate transcription: transcription of any gene in any cell type. Proc Natl Acad Sci USA. 1989; 86: 2617-21.
19. Takemoto CM, Yoon YJ, Fisher DE. The identification and functional characterization of a novel mast cell isoform of the microphthalmia-associated transcription factor. J Biol Chem. 2002;277: 30244-52.
20. Busam KJ, Iversen K, Coplan KC, Jungbluth AA. Analysis of microphthalmia transcription factor expression in normal tissues and tumors, and comparison of its expression with S-100 protein, gp100, and tyrosinase in desmoplastic malignant melanoma. Am J Surg Pathol. 2001; 25: 197-204.